|
|
|
|
|
|
|
| << | < | > | >> |Indice
Prologo 13
PRIMA PARTE
STRUTTURA E MOVIMENTO
1. Segui la corrente 19
2. Ingegneria strutturale 47
3. Imparare a muoversi 71
4. Di testa propria 93
5. Durerà una vita 119
6. Complessità semplice 143
SECONDA PARTE
RILEVAZIONE A DISTANZA
7. Un universo chimico 167
8. Il verso giusto 191
9. Il sonar animale 215
10. Vedere la luce 249
11. L'epica andata e ritorno 277
12. Parlare con ali animali 305
Epilogo 329
Note 331
Per saperne di più 367
Bibliografia 371
Ringraziamenti 389
Indice analitico 391
|
| << | < | > | >> |Pagina 13Tu sei una Grande Scimmia. E si dà il caso che entrambi gli autori siano anch'essi animali appartenenti a questa famiglia, che condividono le rispettive dimore con altri ominidi oltre che con mammiferi della famiglia canina e felina. Senza volerlo, ospitiamo a casa nostra anche una vasta gamma di altri animali, appartenenti per lo più alla classe degli insetti e degli aracnidi. In alternativa avremmo potuto cominciare dichiarando «Tu sei un bipede», per poi parlare, continuando in progressione aritmetica, della condivisione della nostra abitazione con quadrupedi esapodi e ottapodi. Tuttavia, l'aver definito i nostri lettori «scimmie» piuttosto che «bipedi» solleva un problema. Probabilmente qualcuno avrà avuto un soprassalto quell'epiteto essendo l'equivalente metaforico di un ceffone , il che non sarebbe accaduto se vi avessimo chiamati «bipedi». Come mai è così? Sono vere entrambe le affermazioni, eppure noi ominidi della specie Homo sapiens non c'è dubbio che i nostri lettori siano solo di questa specie: sarebbe molto interessante apprendere che il libro viene letto anche da non umani ci riteniamo talvolta superiori al resto del regno animale e ce ne abbiamo a male quando ci rammentano la nostra discendenza. Sotto certi aspetti gli umani sono effettivamente superiori, ma per altri versi non lo sono affatto. Considerando la questione con obiettività scientifica e in una certa prospettiva concreta che più sovente si ritrova nell'ingegneria che nella scienza possiamo valutare fino a che punto gli esseri umani reggono il confronto con altri animali per ciò che riguarda le capacità biologiche. Per quanto ne sappiamo, il nostro cervello è il solo capace di pensiero astratto e di linguaggio, capacità che ci colloca in una classe a sé. La nostra vista è piuttosto buona in confronto al resto del mondo animale, anche se non rappresenta il meglio. Altri sensi sono invece deboli o assenti. I cani storcerebbero il naso, non al nostro odore ma alla nostra capacità di fiutare gli odori, e le nostre capacità uditive i gufi non le starebbero nemmeno a sentire. Il nostro scheletro è una dotazione standard, benché forse non così adattato alla locomozione bipede quanto lo scheletro di molti quadrupedi alla locomozione a quattro zampe; se come noi i polpi fossero capaci di alterigia, ci guarderebbero dall'alto in basso per come manchiamo della minima sensibilità alla luce polarizzata, per non dire della capacità di emetterla; i pipistrelli potrebbero ritenersi al vertice della scala evolutiva essendo forniti di un sonar migliore di quelli a più alta tecnologia in nostro possesso. Questo libro tratta delle meravigliosamente multiformi, sbalorditive abilità degli animali, la cui progettazione biofisica è qui considerata dal punto di vista ingegneristico. Gli scheletri costituiscono meraviglie d'ingegneria così come dell'evoluzione; in altre parole, l'evoluzione ha provvisto gli animali di strutture portanti ingegnerizzate (vale a dire dimensionate e realizzate) correttamente, né può essere altrimenti. Gli uccelli sono adeguatamente ingegnerizzati per il volo. I piccioni sono piattaforme volanti di rilevazione a distanza, con prestazioni che possiamo apprezzare in termini ingegneristici e che comprendono strumentazione per la navigazione celeste, ricevitori acustici a banda larga, ricevitori ottici ad alta risoluzione, sensori di campo magnetico. Una piccola mosca è in grado di individuare con grande precisione le sorgenti acustiche, benché le sue dimensioni siano di gran lunga inferiori alla lunghezza d'onda dei suoni che percepisce: un autentico exploit. Gli albatri coprono lunghissime distanze sopra gli oceani australi con un minimo dispendio di energia: sfruttano il gradiente del vento impiegando una tecnica nota ai piloti di aliante come «veleggiamento dinamico». Questi ultimi due esempi, due piccoli frammenti della vasta ricerca compiuta per scrivere il libro, hanno generato un paio di monografie di carattere didattico, scritte per mostrare agli studenti delle facoltà scientifiche come nel mondo animale si faccia uso di solidi (e innovativi) princìpi dell'ingegneria per conseguire determinati scopi. In questo libro troverete i risultati delle nostre indagini, senza passare per la matematica. Che cosa ci qualifica a scrivere un libro sull'ingegneria degli animali? Be', in primo luogo siamo noi stessi degli animali, e lo siamo da diversi decenni, e poi vantiamo entrambi molti anni di esperienza come ingegneri di Ricerca&Sviluppo, avendo lavorato presso multinazionali aerospaziali nella progettazione di algoritmi radar e sonar in campo militare e per altre applicazioni di rilevazione a distanza. Tutti e due siamo laureati in fisica, e questo connubio scientifico-ingegneristico, al quale si aggiungono dei lunghi comuni trascorsi nella modellazione matematica e al computer dei fenomeni fisici, ci dota di un approccio pragmatico allo studio dei meccanismi naturali. Nell'applicare la nostra esperienza ingegneristica al mondo degli animali abbiamo potuto verificare più e più volte quanto essi siano bene adattati e ben costruiti per i ruoli che svolgono, rappresentando ciò che gli ingegneri chiamerebbero una «tecnologia matura». Esaminare gli animali in questa maniera, analizzarli come se fossero dei prodotti dell'ingegneria, non smorza in alcun modo la nostra sensazione di meraviglia di fronte all'eterogeneità, all'adattabilità, e alle capacità straordinarie che dimostrano. Siamo sicuri che la lettura di questo libro vi darà modo di meglio apprezzare le Grandi Scimmie antropomorfe nonché la nostra, più distante, parentela biologica. | << | < | > | >> |Pagina 42Ma perché noi umani dovremmo essere biologicamente un caso a parte riguardo all'energia consumata per unità di peso nel corso dell'esistenza? Perché siamo animali (relativamente) intelligenti. Ogni grammo di materia grigia richiede circa dieci volte l'energia di un'uguale quantità di muscolo o di altro tessuto organico; quanto la potenza specifica assorbita da un muscolo della gamba di un maratoneta. Ben il 20% del nostro tasso metabolico è dovuto a un cervello affamato di energia, benché costituisca solo il 2,5% del peso corporeo; è dunque evidente che esso deve dare vantaggi in termini di sopravvivenza (scoprire perché è facile: usate la testa), ma il costo è pur sempre considerevole. La massa cerebrale è limitata dalle risorse energetiche disponibili, e il funzionamento del cervello deve essere ottimizzato in funzione della massima efficienza; di conseguenza il flusso di energia influenza il nostro modo di pensare. Non c'è bisogno di approfondire i motivi per cui la materia grigia brucia tanta energia, basta ricordare che tutte le strutture altamente organizzate ne consumano già solamente per autosostentarsi. Pensate a un giocoliere che mantiene in equilibrio dei piatti facendoli roteare in cima a delle pertiche; se ne va intorno mulinando ognuna di esse per non fare cadere i piatti, e più piatti e più pertiche sostiene, più fatica fa. La selezione naturale ha scelto di dotarci di un grande cervello, e ne abbiamo pagato in altro modo il prezzo: tirando la cinghia del bilancio energetico in differenti parti del corpo, che difatti fanno economia di energia per esempio con minore crescita, o ridotto impiego a scopo locomotorio, o con una dieta alimentare di qualità più elevata (la cottura del cibo può alleggerire il costo della digestione, il quale ricorderete nella maggior parte dei mammiferi vale il 20% del bilancio energetico). Ricerche recenti indicano che il cervello dei mammiferi non scala con il peso corporeo secondo la legge di Kleiber: misurazioni particolareggiate rivelano che il metabolismo aumenta con W^0.86, con errore di misura dell'esponente pari a solo 0,03 (Wè qui il peso del cervello e non di tutto il corpo). Se così è, al crescere della grandezza il cervello incrementa il proprio fabbisogno di energia più rapidamente del corpo; può darsi che i neuroni amplino le proprie connessioni o lavorino più sodo in cervelli relativamente grandi. Un matematico direbbe che il vostro cervello ha un'area frattale di dimensione 5 o 6, ma badate a non scervellarvi attorno a questo concetto tanto da mettere a repentaglio il vostro equilibrio energetico. | << | < | > | >> |Pagina 47La struttura di un animale è fondata sul suo scheletro, proprio come quella di un edificio sull'intelaiatura; queste sono estremamente differenziate, e gli scheletri altrettanto. Molti grattacieli posseggono un'anima centrale che sostiene il peso dell'intera costruzione: una spina dorsale. La concezione di altri edifici prevede che siano sorretti dalla stessa muratura esterna: un carapace, o esoscheletro. Differenti progetti soddisfano differenti esigenze, e una delle principali impressioni destate dalla rassegna anche superficiale dell'anatomia animale riguarda la grande varietà strutturale esistente. Una qualunque buona idea, per esempio lo sviluppo di una proteina dura e fibrosa come la cheratina, sboccia nel corso di eoni in centinaia di varianti (corna, peli, lane, unghie, aculei, artigli, zoccoli, scaglie, penne, fanoni, becchi, conchiglie), ognuna precisamente affinata dall'evoluzione per adattarsi a una specialissima funzione. Quest'ampia variabilità rende impegnativo stendere un resoconto didascalico dell'ingegneria delle strutture animali: se lo vogliamo esauriente esso richiederebbe un'enciclopedia in più volumi, se vogliamo limitarci alla panoramica per sommi capi dei princìpi informatori, si corre il rischio di perdere di vista la realtà di tutti i giorni. La difficoltà si aggrava ancora se consideriamo che gli aspetti ingegneristici delle conformazioni animali non stanno solo nello scheletro; esse non sono infatti statiche, richiedono potenza propulsiva, muscoli, e questi a loro volta necessitano di energia e di un sistema circolatorio. E si potrebbe continuare.
L'impostazione che abbiamo adottato in questo capitolo aggira in un
certo senso l'ostacolo: non volendo essere né prolissi né inconsistenti,
né succinti né ampollosi, per riuscire sia formativi che leggibili presentiamo
alcuni concetti e principi ingegneristici basilari ai quali tutte le
strutture (scheletri e sistemi di circolazione in particolare) si uniformano,
inframmezzati da esempi molto specifici, alcuni dei quali estremi.
Dagli uni agli altri ce ne passa, e potreste dovere riflettere un poco per
scorgere il nesso, talvolta crudamente delineato, tra il disegno generale
e la sua implementazione. Per esempio, ai pesci toccherà un ruolo di
primo piano nell'esposizione delle diversità strutturali tra le varie classi
zoologiche; dalla semplice teoria delle travi salteremo alle ossa degli
uccelli; a parlarci della circolazione del sangue saranno le giraffe, previa
breve dissertazione sul moto dei fluidi nelle tubature. Dato l'ampio
spettro di argomenti trattati, in questo più che negli altri capitoli dovrete
affidarvi per colmare le lacune ai riferimenti di bibliografia; qui troverete
è vero delle risposte, ma soprattutto riceverete gli strumenti
per porre più approfonditi interrogativi.
I biologi sospettano che le specie viventi finora scientificamente descritte (1 700 000) non siano che una piccola parte di tutte quelle che esistono. Per la metà si tratta di insetti, essendoci più tipi di coleotteri (290 000) che di vegetali. Questa profusione di insetti non deve sorprendere: un argomento ecologico basato su leggi di scala indica che il numero di specie a sua volta proporzionale al numero di nicchie dovrebbe aumentare al diminuire del peso corporeo (N ∝ W^-1/3), cosicché un gruppo di animali che pesano 1000 volte meno di quelli di un altro gruppo dovrebbe suddividersi in un numero di specie 10 volte più grande (vi sarebbero, per fare un esempio, 10 specie di topo per ciascuna di cervo). Delle 55 000 specie conosciute di vertebrati circa la metà (il 52%) sono pesci. Come mai così tante? Non esistendo nicchie ecologiche in un metro cubo d'acqua, in mare non potremmo certamente fare uso di un'argomentazione di scala, e altrettanto certamente gli habitat di terra sono più diversificati di quelli marini. Non dovremmo quindi attenderci un maggior numero di specie terrestri? In questo paragrafo vogliamo mettere in evidenza la pura e semplice diversità di fattura degli animali, e come strumento esplicativo scegliamo i pesci precisamente perché non è così ovvio il motivo per cui sono tanto diversi tra loro. (Le considerazioni di scala alla fine si applicano anche ai pesci ci sono più specie piccole che grandi , ma pure la storia evolutiva e la geologia giocano una parte di rilievo). Rivolgiamoci innanzitutto ai piani strutturali corporei più elementari, risalenti a un'epoca molto, molto remota della storia dell'evoluzione. Le prime creature erano masse di cellule di forma sferoidale, ma lo sviluppo del tratto digestivo (nella sua forma più semplice, un tubo dalla bocca all'ano) determinò in esse una simmetria assiale: immaginatevi ad esempio una sfera attraversata da un foro. (Di solito una simmetria di questo tipo è definita, impropriamente, «radiale» o «raggiata», ma per un fisico o un ingegnere è la sfera ad essere radialmente simmetrica, mentre forme come un cilindro o una sfera forata lungo un diametro hanno simmetria assiale). Molti animali acquatici mantengono la primitiva simmetria assiale (la medusa è uno di questi), che funziona bene per esseri sedentari verso cui gli stimoli e il cibo possono arrivare, fluttuando, da ogni direzione (fig. 9). D'altra parte creature così lente o immobili vengono facilmente predate, e così molte hanno sviluppato strutture protettive, come aculei, conchiglie, o cellule urticanti. Il passo evolutivo seguente ridusse ulteriormente la simmetria da assiale a bilaterale. La gravità determina un'asimmetria tra parte superiore (dorsale) e parte inferiore (ventrale), ma non tra destra e sinistra. Hanno simmetria bilaterale pesci, uccelli, la maggior parte degli animali terrestri, e tutti i vertebrati compresi voi stessi: la metà destra e sinistra della struttura esteriore, oltre che le parti interne, sono immagini (quasi) speculari. Per la locomozione la simmetria bilaterale è più adatta di quella assiale: è difficile immaginare un pesce nuotare o un uccello volare in sua assenza. La simmetria bilaterale consente all'animale di assumere una forma idro-/aerodinamica. Essa permette anche il posizionamento degli organi interni in parti distanti del corpo; in particolare gli organi di senso possono venire raggruppati frontalmente, dove maggiormente servono. Questa aggregazione sensoriale conduce alla cefalizzazione, vale a dire lo svilupparsi di testa e cervello. Il medesimo principio ingegneristico si applica a ogni macchina che si muova di moto autonomo: un aereo è bilateralmente simmetrico, con la strumentazione di rotta collocata anteriormente, e questa impostazione è basilare al punto che esiste da oltre un secolo, dagli esordi del volo con pilota a bordo (che seguì il breve preambolo dei palloni aerostatici a simmetria assiale). La simmetria bilaterale animale risale a prima ancora: a 500 milioni di anni fa, al periodo Cambriano, nel corso del quale esplose la diversità dei piani strutturali degli animali. Di qui alla segmentazione (un'altra innovazione evolutiva di grande successo) il passo è breve. Gli insetti sono ovviamente segmentati, ma quanto a questo lo siamo anche noi (basti pensare alla colonna vertebrale). La segmentazione è una estensione semplice dal punto di vista algoritmico dell'organizzazione del piano strutturale anatomico. Essa rende più facile la codifica della struttura animale ai fini dello sviluppo, perché le istruzioni sono sempre le stesse, e fra gli altri vantaggi è utile per la locomozione.
Torniamo ora ai nostri pesci. È generalmente riconosciuto che nei pesci esistono sei forme corporee di base, a loro volta ulteriormente suddivise. I predatori sono da inseguimento (come il tonno, che si sposta rapido ed è quindi affusolato, spesso di grosse dimensioni e solitamente a coda forcuta) oppure da agguato (come il luccio, con bocca grande e pinne concentrate dietro per facilitare la brusca accelerazione). I pesci di superficie (per esempio il guppy, Poecilia reticolata) sono piccoli e con la bocca orientata verso l'alto, mentre quelli di fondale (per esempio la passera pianuzza, Platichthys flesus) hanno corpi appiattiti e occhi piccoli. I pesci dal corpo compresso lateralmente (come il pesce luna, Mola mola) di solito hanno grandi le pinne e gli occhi; gli anguilliformi hanno foggia allungata a mo' di fune o nastro, con testa tozza che si raccorda alla sezione trasversale del corpo. Perché tanta varietà di forma e dimensioni? Potremmo aspettarci che gli abitatori dei fondali esibiscano la stessa diversità degli animali terrestri, ma gli altri? Esistono ambienti acquatici di vario tipo: quelli di acqua dolce ospitano un terzo delle specie ittiche; quelli marini di acque poco profonde si dividono in caldi e freddi; infine gli habitat marini di profondità possono essere pelagici (in mare aperto) o bentonici (fondali d'alto mare). Ma queste suddivisioni non sono così nette o così numerose come gli habitat terrestri. Un indizio per arrivare a capire la diversità dei pesci è dato dalle linee generali dello sviluppo evolutivo. I pesci primitivi erano grandi e veloci, con corpi affusolati e pinne raggruppate nella metà posteriore. Le specie successive erano di taglia più piccola e provviste di un maggior numero di aculei, con le pinne distribuite sul corpo con maggiore uniformità. Alla frammentazione, iniziata circa 200 milioni di anni fa, della Pangea il supercontinente che un tempo comprendeva tutte le terre emerse del pianeta seguì una fase di rapido (in termini evoluzionistici) sviluppo. La divisione in continenti più piccoli determinò infatti un aumento della lunghezza complessiva della linea costiera, accompagnato dalla formazione di regioni litoranee e di zone di platea poco profonde, bacini, questi, ricchi di sostanze nutritizie e bene irraggiati dal sole, e dunque con abbondante biomassa. A causa di questa estensione il numero di pesci e delle loro specie si accrebbe astronomicamente. Prima della spaccatura della Pangea la maggior parte della superficie terrestre era costituita da mare aperto, cosicché sia i predatori sia le prede dovevano essere il più possibile veloci. Dopo, i mari poco profondi offrivano scogliere coralline, fondali d'alghe, e altre nicchie e recessi adatti per nascondersi, e i pesci divennero più piccoli per sfruttare queste nuove opportunità. Alle prede e ai predatori serviva manovrabilità più che velocità pura, e le pinne si ridistribuirono. E quando la rapidità venne a mancare, come metodo di autodifesa si svilupparono gli aculei. In conclusione, la grande varietà dei pesci (fig. 10) mostra che la forma generale della struttura animale dipende sì dall'ambiente e dalle abitudini, ma anche dal capriccio dei trascorsi evolutivi e dei cambiamenti geologici. | << | < | > | >> |Pagina 53La grande maggioranza, per non dire tutti i lettori di questo libro possiedono un endoscheletro, la cui funzione principale è di fornire il sostegno meccanico. Ogni osso deve così essere abbastanza robusto da sopportare le forze e i momenti che prevedibilmente agiranno su di esso, e deve connettersi alle altre ossa in maniera tale che il suo orgoglioso possessore possa muoversi liberamente. Il complesso dimensionamento di uno scheletro è di conseguenza in larga parte un problema di ingegneria, al quale possiamo ragionevolmente applicare le nostre nozioni di meccanica per ricavare indicazioni su come gli scheletri siano costituiti. Per questo non avremo bisogno di addentrarci estensivamente nella matematica dell'analisi degli sforzi: ci basterà citare una sola formula per spiegare come gli ingegneri descrivano le sollecitazioni meccaniche e come essa si rapporti alla biomeccanica dello scheletro. La useremo per distinguere quali aspetti del dimensionamento scheletrico noi progettisti possiamo modificare a piacimento e quali invece sono fissi per motivi diversi dall'analisi delle sollecitazioni. Sottoposti a sforzo assiale per sostenere un peso, una colonna architettonica in pietra o un pilastro d'acciaio possono collassare se il carico è sufficientemente elevato. L'equazione del calcolo strutturale che stabilisce il massimo carico di punta sostenibile da un'asta è la classica formula di Eulero: Per sollecitazioni di tipo diverso, come flessione o torsione, ci sono altre formule. Nella formula, Fcr è il carico critico, L la lunghezza dell'elemento e k una costante adimensionale il cui valore dipende da come è incastrato ai due estremi (per esempio, se è saldamente infisso nel terreno il valore è maggiore, se è solamente incernierato è minore). E è il modulo di Young, una costante che descrive la rigidità intrinseca del materiale. La pietra è più rigida della pasta sfoglia, e quindi ha un E molto più grande, e questa è una delle ragioni per cui le colonne del Partenone sono fatte di pietra e non di pasta sfoglia. Il termine restante della formula è I, un parametro geometrico che va sotto la sgraziata (e, per un fisico, sconcertante) denominazione di «momento d'inerzia della sezione trasversale», anche noto come «momento di superficie del secondo ordine»; esso descrive il modo in cui la sagoma della sezione (quadrata o circolare, cava oppure piena, e così via) influisce sulla resistenza della struttura. Sostituendo nella formula i valori appropriati dei parametri, un progettista è in grado di capire ad esempio se una data colonna reggerà un dato carico; in caso negativo potrà aumentarne la robustezza accorciandola, incastrandola in modo diverso, impiegando un altro materiale, oppure variandone la forma. Ma non dovrete sgobbare con la matematica; non intendiamo mettervi alla prova sulle astruse denominazioni delle varie grandezze o bersagliarvi con le tante formule dell'ingegneria meccanica che forniscono la resistenza di una trave o di un pilastro assoggettato a sforzo di tensione, compressione, taglio o torsione. La formula di Eulero viene menzionata solamente per mostrare come mettere a frutto l'analisi strutturale nella descrizione degli scheletri (è chiaro oramai che le ossa della gamba si possono considerare come colonne che sorreggono un carico). Dalla formula si vede ad esempio che le ossa lunghe sono soggette a instabilità sotto carico di punta (ovvero a collasso per incurvamento) più di quelle corte, un fatto che il lettore può verificare sperimentando con uno spaghetto crudo e asciutto. Sulle ossa dello scheletro agiscono forze diverse, e questo si rispecchia nella loro struttura. Le ossa sono per esempio connesse tra loro in vari modi: all'articolazione sferoidale (o enartrosi) della spalla o dell'anca è associato un valore di k diverso da quello dell'articolazione a cerniera del gomito o del ginocchio. Si tratta di vincoli di natura funzionale, più che meccanica: il giunto a sfera consente più grande libertà di movimento, mentre quello a cerniera sopporta un carico maggiore. Anche la lunghezza delle ossa dipende dalla funzione; assumiamo quindi che per un determinato scheletro i termini k e L nella formula di Eulero siano invarianti. Il modulo di Young delle ossa è molto alto, il che significa che l'osso è molto rigido. Certo non ha la rigidità dell'acciaio, ma i sistemi biologici non si sono ancora evoluti al punto che gli scheletri di acciaio si sviluppino naturalmente o forse il peso dell'acciaio lo rende meno vantaggioso dell'osso. La natura ha scelto l'osso perché è il materiale con elevato modulo di Young che può produrre più rapidamente. Anche E è dunque fissato. La forma delle ossa, rappresentata nella formula di Eulero dal termine I, è qui per noi del più grande interesse. Avendo visto che tutti gli altri parametri sono vincolati, dobbiamo ora domandarci: che forma deve avere un osso? Supponendo che il peso dell'osso sia predeterminato in altri termini, che la natura abbia assegnato a ogni osso una certa quantità di materiale , come dovrà essere sagomato questo materiale per avere la massima resistenza? Questa è una questione di ingegneria, non di funzione. La funzione di un osso determina il tipo e l'intensità delle forze che agiscono su di esso, così come la sua lunghezza e il tipo di connessione con altre ossa, ma trovare la forma migliore di quel particolare osso è essenzialmente un problema ingegneristico.
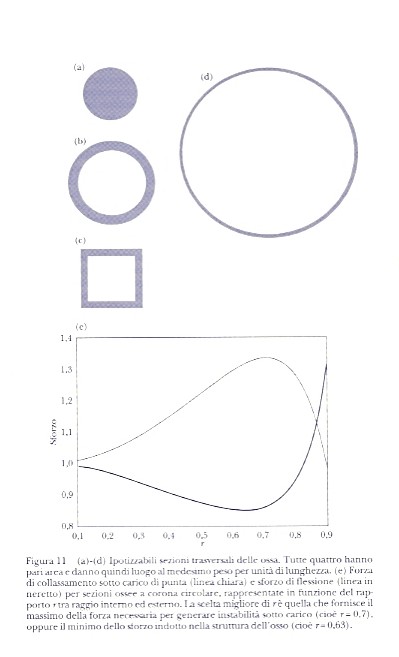
Consideriamo la sezione trasversale di un osso. Nella figura 11 viene avanzata la candidatura di una serie di possibili sezioni; tutte hanno la stessa area, e quindi daranno luogo allo stesso peso per unità di lunghezza. E il peso è un aspetto importante, in quanto un osso leggero per essere spostato richiede meno energia di uno più pesante; in compenso un osso leggero è più fragile. Serve quindi un compromesso tra peso e resistenza. Le ossa mostrate nella figura possiedono una resistenza all'instabilità sotto carico molto diversa, perché molto diversi sono nei quattro casi i valori di I nella formula di Eulero. Potreste pensare che un osso pieno sia più robusto di uno cavo di pari raggio, e avreste ragione; tuttavia la sezione cava della figura 11b resiste all'instabilità o alla flessione quattro volte più di quella piena della figura 11a (lo sappiamo dal calcolo del momento d'inerzia I delle due sagome), e la sezione ossea della figura 11d venti volte tanto. Qual è dunque il modello migliore? Sappiamo tutti che le grandi ossa dei mammiferi sono cave, cosicché la migliore alternativa dev'essere questa: le ricerche dimostrano infatti che le lunghe ossa cave dei mammiferi sono ottimizzate per il massimo rapporto tra resistenza a flessione e peso. La sezione normale ottimale è una corona circolare con raggio interno pari al 60% di quello esterno, risultato ricavato sulla scorta di più fattori interessanti la progettazione strutturale e non del solo momento d'inerzia geometrico. Le ossa cave contengono il midollo, il cui peso va tenuto in considerazione nel calcolo del peso complessivo. Alle ossa sono attaccati i muscoli; se esse sono deboli richiedono più muscoli, e dunque anche la massa muscolare potrebbe essere un fattore da tenere presente. Possiamo provare che è sufficiente considerare il momento d'inerzia della sezione normale e la densità del midollo (trascurando il più complicato effetto dato dai muscoli) per giungere a una forma ossea ottimale. Il risultato è un po' diverso da quello fornito dalle ricerche più dettagliate, ma non poi tanto. Considerando dunque solo I e tenendo conto della massa del midollo abbiamo calcolato le curve riportate nella figura 11e, che rappresentano la forza di collassamento sotto carico di punta, e lo sforzo di flessione, in funzione del rapporto r tra raggio interno ed esterno della sezione a corona. Si vede che il valore massimo della forza di collassamento interviene a r=0,7, e il minimo dello sforzo di flessione attorno a r=0,63. Un calcolo semplificato mostra che ad altre sollecitazioni, ad esempio la torsione, resiste meglio la sezione piena. Tuttavia nei casi d'interesse pratico gli sforzi di torsione sulle ossa si verificano di rado o sono trascurabili rispetto agli sforzi di flessione, per cui la natura non ne ha tenuto conto, mentre tiene conto dei secondi. Il valore ottimale calcolato si avvicina al nostro valore medio di r per le ossa vere (e con i calcoli completi e dettagliati si va ancor più vicino). Come si vede, le forme in figura 11 non rappresentano l'optimum: il modello della figura 11b è quello che più vi si avvicina, anche se lo spessore ottimale per un osso è quasi il doppio di questo. Nonostante le loro limitazioni, le forme mostrate in figura 11a e 11d compaiono entrambe in natura, poiché il rapporto resistenza-peso non sempre è il fattore decisivo. Le ossa piccole tendono ad essere piene, dato che il loro peso è comunque basso e l'osso pieno è più resistente; in questi casi la robustezza prevale sul rapporto resistenza-peso. Quando invece è il peso il fattore determinante troviamo ossa cave e sottili, come nella figura 11d. Lo pteranodonte (Pteranodon), un dinosauro alato gigante di 80 milioni di anni fa con un'apertura alare di 9 m, necessitava per il volo di ossa lunghe e rigide, e al tempo stesso estremamente leggere; le ricostruzioni indicano che le ossa di pteranodonte avevano in effetti grande diametro e piccolissimo spessore. Il prezzo da pagare è che ossa di questo tipo sono per altri versi deboli. Per esempio, l'osso mostrato nella figura 11d verrebbe frantumato facilmente dalle zanne di un predatore. E che dire della sezione ossea della figura 11c? Sembra che la natura abbia trascurato la possibilità di una forma quadrata (fatta forse eccezione per il favoloso C. ridiculus del capitolo 1). Le sagome circolari funzionano meglio, perché sono egualmente resistenti in ogni direzione. Un carico di punta deve necessariamente applicarsi in senso longitudinale all'osso, ma una forza flettente giace su un piano trasversale e può avere un angolo qualunque. Il problema di un osso quadrato è che la sua resistenza all'incurvamento non sarebbe costante, dipendendo dall'angolo di applicazione della forza. Se la sollecitazione è verticale o orizzontale, l'osso quadrato della figura 11c è resistente quanto quello della figura 11b; con altre angolazioni invece è più debole (il tallone d'Achille di una sezione quadrata è negli angoli, dove lo sforzo è maggiormente concentrato e dove più facilmente si verifica la rottura). Ovviamente in natura gli sforzi di flessione (come di ogni altra forza) possono avere una direzione qualsiasi: un animale che lotta, o per effetto di una caduta, deve attendersi colpi da ogni parte. Ecco quindi che i cerchi hanno la meglio sui quadrati. | << | < | > | >> |Pagina 81
Dato che il volo degli uccelli è assai più complesso di quello degli aeroplani, le analisi tecniche nei due casi sono sovrapponibili, ben che vada, solo parzialmente. Possiamo utilizzare almeno alcune delle notevoli conoscenze acquisite dagli ingegneri aeronautici sull'aerodinamica dell'ala fissa? In base alla seguente argomentazione la risposta è affermativa. La velocità di sbattimento delle ali degli uccelli in volo veloce è molto inferiore alla velocità con cui l'uccello si sposta nell'aria, cosicché appare ragionevole modellare aerodinamicamente quei pennuti come aeromobili ad ala fissa, a patto di considerare i risultati come approssimazioni (se l'ala si muove molto poco nel tempo impiegato dall'uccello a coprire una distanza pari alla lunghezza del proprio corpo, possiamo considerarla approssimativamente come fissa). Il criterio non vale per uccelli in volo stazionario, meglio modellati mediante l'aerodinamica dell'elicottero. In entrambi i casi procederemo con cautela, e vedremo a quali risultati conducono tali studi ingegneristici.
La figura 19a illustra la fisica di base del volo degli uccelli. I muscoli azionano le ali, il cui movimento genera la spinta F (la forza che proietta l'uccello in avanti) e la portanza L (lift, la forza che lo sostiene). A queste forze si oppongono rispettivamente la resistenza aerodinamica D (drag) e la forza peso W; e fin qui, tutto bene. Considerazioni di maggior dettaglio mostrano che la potenza sviluppata da un volatile dovrebbe in teoria avere andamento simile al grafico della figura 19b. La curva si impenna a velocità elevata, perché la resistenza aerodinamica cresce più che linearmente con la velocità, e occorre maggiore potenza per vincerla. Ma aumenta pure a bassa velocità, perché il volatile che si sposta lentamente ha difficoltà a produrre portanza; basta pensare a quanto energicamente deve battere le ali il gheppio (Falco tinnunculus) mentre se ne sta sospeso in aria. La forma generale della curva mostra quindi che esiste una velocità ottimale v1 per la quale la potenza impiegata è minima. Quando vola a quella velocità, un uccello consuma meno energia. Per ridurre al minimo il dispendio energetico richiesto dal volo, la velocità di un uccello non può quindi discostarsi da certi valori. Ovviamente la forma dettagliata della curva di potenza cambia da una specie all'altra, ma sembra proprio che molti uccelli volino di preferenza a certe velocità, corrispondenti più o meno al minimo dispendio. Esiste però una seconda velocità ottimale, contrassegnata come v2 in figura 19b. Il volatile potrebbe non cercare di volare alla minima energia possibile, se per lui fosse maggiormente importante giungere quanto più lontano possibile (come durante una migrazione). Questa velocità cui corrisponde la massima autonomia è paragonabile al passo che adotta un maratoneta: è la velocità da tenere per coprire la più lunga distanza con un pieno, e non coincide con la velocità meno dispendiosa (alla velocità v1 il nostro volatore spende la minima energia per unità di velocità, alla velocità v2 spende la minima energia per unità di distanza). Si ricava dal calcolo che v2 è in corrispondenza del punto dove la retta passante per l'origine è tangente alla curva, come indicato dalla linea tratteggiata nella figura. Anche di queste conclusioni sussistono riscontri sperimentali: gli uccelli migratori effettivamente volano a velocità prossime al valore teorico v2 proprio della loro specie. Un altro risultato generale che viene dall'aerodinamica e che più o meno si ritrova in natura riguarda la portanza L (fig. 19a) che, per un uccello in volo orizzontale, dev'essere uguale e contraria alla forza peso, altrimenti il volatile guadagnerebbe o perderebbe quota. L'aerodinamica ci dice poi che la portanza è proporzionale all'area A della superficie alare e al quadrato della velocità: L ∝ A v^2. Combinando le due osservazioni con considerazioni geometriche del tutto simili a quelle fatte nel capitolo 1 riguardo alla legge di Rubner, troviamo che la velocità del volo d'uccello scala col peso secondo W^1/6, cosicché è lecito attendersi che una cicogna di 2 kg voli circa due volte più veloce di un passero di 30 g. Calcoli di scala più precisi portano a un esponente un poco diverso, per cui sarebbe v ∝ W^0.18. La maggior parte degli uccelli (e dei velivoli) pressappoco si adeguano a questa relazione di scala, ma non i colibrì e gli insetti quando si librano sospesi, perché i volatori stazionari (o «a punto fisso») non sono paragonabili a piccoli aeromobili ad ala fissa, dato che sono piuttosto minuscoli elicotteri. (Possono, e devono, volare a punto fisso per via del loro peso ridotto che, come abbiamo visto, implica bassa velocità in volo). Come si ricorderà, abbiamo distinto vari tipi di locomozione terrestre in base al valore di un parametro adimensionale (il numero di Froude). Per il volo battente esiste un parametro analogo, il rapporto di avanzamento J, definito da dove v è la velocità relativa all'aria, Φ l'ampiezza della battuta d'ala, f la frequenza di battuta, e l la lunghezza dell'ala. Il rapporto di avanzamento è dunque proporzionale al rapporto tra la velocità di volo, v, e la velocità dell'estremità alare, Φfl. Uccelli e insetti sono dinamicamente simili se volano con lo stesso rapporto di avanzamento, e così ci aspettiamo che un insetto che vola lento batta le ali a frequenza maggiore di un uccello che procede lentamente, il quale ha ali più lunghe. Il parametro che abbiamo introdotto consente di ricavare un'altra relazione di scala. Poiché J non ha dimensioni, e poiché v scala più o meno con W^1/6 e l con W^1/3, a parità di Φ dall'espressione di J risulta subito v ∝ W^(1/6-1/3) = W^-1/6; i volatili più pesanti devono dunque sbattere le ali più lentamente di quelli più leggeri, e ancora una volta si osserva in natura che la regola è in linea di massima verificata. Una notevole complicazione che emerge quando si tenta un'analisi quantitativa del volo battente sta nell'estrema diversificazione del fenomeno: ci riferiamo non solamente alla grande varietà di uccelli con tutte le loro forme e dimensioni, ma anche alle disparate modalità del volo battente. Il battito alare non è normalmente continuo: in molti piccoli uccelli sono presenti interruzioni del battito che avvengono in modi diversi e portano a distinguere tre regimi differenti. Nel volo «intermittente» si susseguono un periodo di sbattimento ali, durante il quale il volatile guadagna spinta e portanza, e uno in cui le mantiene ripiegate lungo il corpo e, così diminuendo la resistenza, fora l'aria come un proiettile. Vista di lato, la traiettoria nel volo intermittente appare sinusoidale, e la quota di volo dell'uccello si alza e si abbassa periodicamente. Molti uccelli piccoli e dalle ali corte, ad esempio i passeri, prediligono questa modalità. Il volo «ondulato» esibisce invece una fase remigante seguita da una di planata; è più diffuso del primo e lo adottano uccelli con ali più lunghe, quali corvidi, piciformi e rapaci; però un volo di questo tipo è pure praticato dallo storno comune (Sturnus vulgaris), nonostante abbia ali corte. La terza, meno comune condotta in volo è differente, nel senso che non comprende una fase di riposo. Al contrario, il volo «sfarfallato», come quello della gazza (Pica pica), consiste nello sbattimento alare continuo ma a due frequenze differenti, che si alternano. La modellazione dettagliata di questi tre tipi di volo alternato indicherebbe un risparmio di energia di circa il 10%. | << | < | > | >> |Pagina 86Siamo in grado di analizzare il volo degli uccelli velieri con maggiore sicurezza e precisione di quanto siamo riusciti con il volo ad ala battente, perché veleggiamento e planata sono fattispecie ad ala fissa, che gli ingegneri aerospaziali comprendono piuttosto bene. Gli albatri si sono evoluti per planare e veleggiare, e anche se aquile ed avvoltoi sono anch'essi planatori e veleggiatori molto abili, gli albatri sono i migliori; prima spiegheremo il come e poi il perché. La ventina di specie di albatri trovano tutte quante il proprio habitat negli oceani australi, in maggioranza lungo i «Quaranta Ruggenti» e i «Cinquanta Urlanti», le latitudini dove i venti imperversano la gran parte dell'anno, con poche terre emerse in grado di ostacolarli. Questi volatili nidificano su delle isolette e vagano in lungo e in largo a cercare il cibo per sé e per la prole; devono così percorrere grandi distanze, tanto che una spedizione di approvvigionamento può coprire mille chilometri al giorno e durare una decina di giorni. Battere le ali su tali distanze richiederebbe di gran lunga troppa energia, e allora gli albatri si librano in volo e planano. Questa modalità richiede poco o nulla in termini energetici; e difatti vedremo che questi uccelli sono capaci di sfruttare il vento in modo da viaggiare senza alcun dispendio meccanico (e con solo poca energia fisiologica). Come riescono a volare così? Sono in sommo grado bene adattati per il volo a vela e hanno sviluppato una tecnica di «veleggiamento dinamico» che sfrutta al massimo la forza del vento. Gli adattamenti comprendono grandi dimensioni (le ali dell'albatro urlatore, Diomedea exulans, con i loro tre metri e mezzo di apertura sono le più lunghe fra tutti gli uccelli esistenti), forte allungamento alare come in un aliante (chiaramente visibile in fig. 20), e una particolare, unica articolazione della spalla, dove un fascio tendineo incernierato alla scapola blocca le lunghe ali consentendo di mantenerle distese col minimo sforzo. Passiamo ora al nucleo ingegneristico del volo degli albatri: il volo planato e il veleggiamento. In planata, si galleggia sul vento, cedendo energia potenziale per la maggiore distanza orizzontale possibile consentita dalle ali; l'angolo di planata θ è dato dal rapporto tra la resistenza aerodinamica D e la portanza L: tgθ = D/L, cosicché un angolo piccolo (portanza elevata) permette di restare in aria, a partire da una data altitudine, per una lunga distanza. Questi uccelli possono arrivare a un angolo di planata quasi altrettanto ridotto di quello di alianti ben progettati costruiti dall'uomo: circa 3°. Il veleggiamento dinamico è una tecnica di volo che sfrutta il gradiente di vento per estrarne energia. I volovelisti l'hanno imparata: spiraleggiando, come descritto oltre, a ridosso dei rilievi dove la velocità dell'aria varia con la quota, bassa nell'ombra di vento di una parete e alta sopra di essa , l'aliante si libra verso l'alto, acquistando quota facendosi dare un passaggio dal vento. Ma sulle distese oceaniche non ci sono montagne; come riescono allora gli albatri a destreggiarsi nel veleggiamento dinamico? Vicino al pelo dell'acqua, dove l'attrito riduce la velocità del vento, esiste in effetti il gradiente: la velocità del vento acquista così un profilo verticale, passando da zero alla superficie fino al valore massimo a circa venti metri di quota. Lo scienziato ottocentesco Lord Rayleigh avanzò per primo l'ipotesi che gli albatri utilizzino il profilo verticale di velocità della corrente d'aria. Impennandosi contro il vento e scendendo in picchiata in suo favore è possibile come mostrano i calcoli ricavarne energia e coprire lunghe distanze con piccolo sforzo: è questo il veleggiamento dinamico. Questi uccelli sfruttano anche altri e più complessi fenomeni aerodinamici con il discendere fino al livello dell'acqua (così vicini che con la punta delle ali arrivano a toccarla). L'aria spostata dal loro movimento rimbalza dalla superficie fornendo portanza aggiuntiva, ed è questo l'«effetto suolo»; inoltre le onde oceaniche fanno muovere l'aria in loro prossimità in modi che un esperto pilota di aliante, o un albatro, sanno sfruttare, un effetto definibile in questo caso come «aerosostentazione ondosa». | << | < | > | >> |Pagina 167Gli esseri umani sono animali fortemente orientati alla visualità. Facciamo esperienza del mondo attraverso gli occhi, e la vista è talmente preponderante nella nostra vita che le metafore visive dominano il linguaggio. Se non balza agli occhi pure a voi, metteremo ancor più in luce questo punto con un'affermazione diretta: gli esseri umani «vedono» il mondo che li circonda.
Per altri animali non è così; essi si affidano principalmente a un senso
chimico che chiamiamo olfatto; serve loro per individuare le sostanze
nutritive che forniscono energia e la materia prima per la sopravvivenza
e la crescita; serve loro per identificare, ed evitare, i pericoli per la loro
esistenza rappresentati dai predatori o da veleni; serve loro per rintracciare
la fonte di quelle molecole che promettono una possibilità di riprodursi. Non
deve dunque sorprendere che i sensi chimici siano stati i
primi a evolversi e che per molte creature siano ancora i più importanti.
Nonostante esistano animali naturalmente ciechi e naturalmente sordi,
non si conoscono animali che manchino di una qualche sensibilità chimica. Ecco
dunque l'importanza dell'olfatto, ed ecco perché appare davvero strano che
nell'ambito della rilevazione chimica l'uomo sia alquanto inetto.
Più o meno tre miliardi e mezzo di anni or sono negli oceani si formò la vita. Quelle primitive forme viventi avevano bisogno di mezzi semplici per rilevare le sostanza disciolte nell'acqua o «in soluzione», come si dice tecnicamente. Vediamo ancor oggi questo processo al lavoro negli organismi elementari come i batteri, i quali, benché siano creature unicellulari con limitate capacità sensorie, mostrano un comportamento chiamato chemiotassi (o chemotassi) che li spinge verso il nutrimento e lontano dalle tossine. Il batterio possiede, disposti attorno alla membrana esterna della sua unica cellula, svariati tipi di recettori in forma di molecole proteiche capaci di legarsi ad altre sostanze presenti nell'ambiente. Si tratta in pratica di una serie di chiavi e serrature: una particolare chiave chimica entra in un solo recettore-serratura che invia allora un segnale specifico al flagello del batterio, un propulsore filamentoso rotante collocato all'esterno della membrana e per mezzo del quale il batterio può spostarsi. L'attivazione di un recettore legato a un segnale chimico associato al cibo costituito di solito da molecole zuccherine spinge il batterio verso la sua fonte; l'attivazione di un recettore associato a tossine lo spinge invece ad allontanarsene. I batteri possono avvertire anche i gradienti di concentrazione, e avanzare o retrocedere rispetto alla direzione del segnale chimico. In un essere unicellulare privo di mezzi evidenti per il simultaneo confronto del livello di concentrazione chimica in differenti parti del suo corpo, questa capacità implica un qualche cosa di affine alla memoria: il batterio sa che in quel punto la concentrazione è diversa da quella che era nella posizione precedente e di conseguenza continua a seguire la giusta direzione. Come vedremo, questo meccanismo a chiave e serratura costituisce il fondamento di ogni rilevazione chimica animale. Dal semplice batterio al naso sensibilissimo del cane, i sottostanti princìpi fisici non variano; solamente, Fido ha molti più recettori e fa molte più cose con il segnale chimico. | << | < | > | >> |Pagina 172Può sembrar strano affermare che l'olfatto è il principe dei sensi, quando gli esseri umani risultano al riguardo abbastanza inetti ed esso gioca un ruolo abbastanza secondario nella nostra esistenza. Tuttavia e a rischio di generalizzare, poiché il regno animale è un territorio assai esteso per la maggioranza degli altri animali è il senso più importante, utilizzato per esplorare il mondo e comunicare con le altre creature. L'olfatto funziona anche al buio, a distanza, e richiede soltanto un hardware spartano e privo di parti in movimento. In termini scientifici, l'olfatto non necessita che di una proteina recettrice sul contorno di una cellula, che chimicamente si leghi a una molecola esterna con la quale abbia affinità, inducendo un neurone a generare un potenziale di azione. Detto più semplicemente, abbiamo una chiave chimica che si adatta a una specifica serratura e vi innesca una reazione, come per i batteri. Nei vertebrati odore è ciò che si verifica nel naso, o, più esattamente, nella cavità nasale. Tradendo le primitive origini, l'apparato olfattivo individua le sostanze chimiche in soluzione (ad esempio in acqua). Nel caso degli animali acquatici queste sostanza sono già disciolte nel mezzo in cui vivono. Per quelli che respirano in aria le cose sono un po' più complicate: le molecole da riconoscere devono innanzitutto essere volatili in modo da venire respirate assieme all'aria, ma poi vanno anche disciolte in un liquido perché i recettori olfattivi possano legarvisi chimicamente. È una delle ragioni per le quali i vertebrati che respirano in aria tendono ad avere il naso che cola: è il muco delle vie nasali che discioglie le sostanze odoranti preparandole a venir percepite. (Ha anche altre funzioni, quali filtrare i microbi e altri detriti, e per questo anche i pesci hanno il muco nasale). | << | < | > | >> |Pagina 191I trasduttori bioacustici passivi («orecchie») si sono evoluti in molte forme e dimensioni in molte classi di animali; un modo ricercato per dire che l'udito è utile. In questo capitolo condivideremo con voi alcuni concetti ingegneristici associati alla rilevazione acustica passiva (l'ascoltare che cosa c'è là fuori), in quello seguente indagheremo il più complicato mondo della rilevazione attiva (ecolocalizzazione). Qui ci limitiamo alla propagazione del suono nei fluidi aria o acqua e parleremo quindi poco delle vibrazioni che si trasmettono attraverso la materia solida. Anche se si tratta sempre di onde sonore, i recettori che gli animali hanno sviluppato per percepirle sono differenti. Negli insetti, i meccanocettori sensibili alle vibrazioni nei solidi sono spesso strutture adattate in modo specifico, mentre gli uccelli e gli esseri umani avvertono le vibrazioni servendosi di piedi o mani. I ragni hanno una conformazione delle zampe specializzata per captare le più minute vibrazioni delle loro tele, che magari denunciano la presenza di una preda. I monotremi quali l'ornitorinco (Ornithorhynchus anatinus) hanno meccanocettori specializzati; ritorneremo a questi bizzarri animali e ai loro dispositivi nel capitolo 10, mentre qui ci occupiamo senz'altro delle onde sonore nel mezzo aereo e acquatico. | << | < | > | >> |Pagina 195Per molti motivi la capacità di distinguere i suoni è un'ottima cosa, e sviluppare una apparecchiatura adeguata contribuisce alla prosperità collettiva (vedi l'esempio delle api). Ecco di seguito un elenco di vantaggi legati all'udito per mezzo di «protesi acustiche», alcune ovvie altre più sofisticate che esamineremo nel corso di questo capitolo e dei successivi. Comunicazione con conspecifici. Le frequenze emesse da un animale nel cantare, chiamare, ululare, vagire, fischiare, nitrire, grugnire, abbaiare, starnazzare, ronzare, trillare, gridare, coincidono in buona parte con quelle del suo campo uditivo. La conclusione ovvia che se ne trae è che l'udito si sia evoluto in tutte le specie o quasi come ausilio alla comunicazione; a questo riguardo si dice di più nel capitolo 12. Scoprire la preda. Molti predatori ne avvertono la presenza dai suoni che questa emette, intenzionalmente o meno. Vedremo come gufi e civette abbiano affinato in sommo grado questa capacità. - Scoprire i predatori. I conigli, i cervi e molte altre specie-preda che sfuggono ai predatori con la fuga hanno un udito eccellente, che agisce da sistema di preallarme consentendo loro un vantaggio iniziale. Sentire l'allarme. Gli animali sociali dispongono di sentinelle che montano la guardia e avvertono il branco o lo stormo con un richiamo della presenza di un pericolo. - Localizzazione. È chiaramente utile per un animale essere in grado di individuare la fonte di molti tipi di suoni, sia che provengano da possibili predatori, prede, o partner sessuali, sia che si tratti di rumori ambientali (per esempio acqua corrente). Navigazione. Vi sono sempre maggiori prove che alcuni uccelli (per esempio i piccioni) sono in grado di udire infrasuoni a frequenza bassissima prodotti dall'ambiente, e che questa facoltà è di ausilio alla navigazione nei voli a lunga distanza, come le migrazioni. Tratteremo in dettaglio questo argomento nel capitolo 11.
Ecolocalizzazione.
Il sonar biologico è oggetto del capitolo 9, ma è
ovvio che per servirsene l'animale deve innanzitutto poter udire.
Dopo aver eseguito test audiometrici di ogni tipo su ogni specie di creature striscianti, volanti, natanti e serpeggianti, i biologi sono giunti ad alcune conclusioni generali. I mammiferi sono diversi da quasi tutti gli altri vertebrati perché possono udire suoni a frequenza superiore a 10 kHz. I pesci, gli anfibi e genericamente i rettili sono insensibili da 5 kHz in su. Le rane (Anura) sono i soli anfibi bene adattati alla detezione dei suoni trasmessi in aria. Gli uccelli odono piuttosto bene, anche se non tanto quanto i mammiferi, e in un campo di frequenze più limitato. Molti insetti hanno sviluppato una speciale sensibilità uditiva su una o più bande ristrette di frequenze. Il grillo (Gryllidae) ad esempio è sensibile non soltanto alle frequenze dei richiami canori della propria specie, ma anche a una banda di frequenze assai più alte (ultrasoniche) parecchio distanti dalla prime , dove resta in ascolto dei segnali di ecolocalizzazione emessi dai suoi predatori, i pipistrelli. Gli animali più piccoli sono in linea di massima sensibili a frequenze più alte di quelle percepite dagli animali di maggiori dimensioni. Molte specie fanno uso di frequenze all'estremo superiore del loro campo di udibilità per individuare la direzione di una sorgente sonora, e la precisione con la quale riescono a stimarla è variabile. Gli esseri umani percepiscono l'altezza relativa dei suoni (riusciamo a distinguere la differenza di altezza), mentre gli uccelli avvertono l'altezza assoluta (sono direttamente sensibili al numero di cicli al secondo); essi sono anche in grado di percepire molto meglio di noi i rapidi cambiamenti di intensità di un'onda sonora. I mammiferi esibiscono una variabilità enorme (su ben nove ottave) della minima frequenza udibile. La forma dell'orecchio esterno dei mammiferi è estremamente variabile. Quelli che fanno vita fossoria non odono molto bene (ma avvertono le vibrazioni del substrato). Alcuni animali (strigiformi, pipistrelli, odontoceti) hanno evoluto capacità uditive che vanno ben oltre le nostre possibilità sia biologiche sia tecnologiche.
| << | < | > | >> |Pagina 215I più sanno che la tecnologia SONAR (SOund NAvigation and Ranging, «navigazione e misurazione di distanza per mezzo del suono») è impiegata sulle navi da guerra per rivelare la presenza di sottomarini nemici nelle tenebrose profondità marine dove la visione è impossibile. Ritengono (correttamente) che il sonar sia simile al radar, se non che invece di emettere onde elettromagnetiche si serve di onde acustiche; e in effetti il sonar è per molti versi esattamente come il radar, essendo le differenze tra le due tecniche di rilevazione a distanza quasi interamente dovute alla diversa natura delle onde che vengono trasmesse. Le onde sonore differiscono parecchio da quelle elettromagnetiche, e ciò pone problemi differenti all'implementazione pratica delle apparecchiature dell'uno e dell'altro tipo; l'identità di fondo è comunque questa: tanto il sonar che il radar inviano onde verso l'ambiente esterno e ne ricevono l'eco. I segnali di ritorno subiscono poi una quantità di ingegnose manipolazioni (molti algoritmi radar e sonar sono gli stessi) per ricavarne dati sull'obiettivo: distanza, direzione, velocità e natura. Gli animali non hanno radar, e in alternativa impiegano la rilevazione passiva a distanza di tipo ottico (in parole povere, vedono), che costituisce l'oggetto del prossimo capitolo. La gran parte di loro non ha sviluppato metodi per trasmettere onde elettromagnetiche di assegnate forma e lunghezza d'onda quindi niente radar , ma alcuni hanno appreso a fare la stessa cosa con il suono; gli esseri umani parlano, le balene cantano, gli uccelli cinguettano e i pipistrelli ecolocalizzano (e lo fanno, come vedremo, anche molte balene e alcuni uccelli). Il termine «ecolocalizzazione» (o talvolta «biosonar»), adoperato per indicare la tecnica del sonar quand'è utilizzata dagli animali, è stato coniato da Donald Griffin, che attorno agli anni Cinquanta fu il primo a comprendere che i pipistrelli si servono della rilevazione acustica a distanza. I microchirotteri (Microchiroptera) sono il meglio nel mondo animale per quanto riguarda l'ecolocalizzazione, gli odontoceti seguono a ruota, e due specie di uccelli il guaciaro (Steatornis caripensis) e la salangana delle grotte (Collocalia linchi) sono terzi con un buon distacco. Su una scala di raffinatezza dell'elaborazione di segnale i dispositivi sonar progettati dall'uomo si collocherebbero molto prima degli uccelli e qualche lunghezza dopo pipistrelli e balene. In questo capitolo esponiamo i fondamenti della rilevazione ecometrica, di lì espandendo la materia fino alle nostre cognizioni sullo stato dell'arte dei processi tecnologici. Una tale impostazione richiede che, una volta riassunti i rudimenti della rilevazione a distanza, si passino in rassegna gli animali ecolocalizzatori in ordine di prestazioni crescenti, dalla rozza elaborazione di fascia bassa di alcuni volatili, alle assai più notevoli abilità di delfini e loro confratelli, fino agli sbalorditivi e non ancora del tutto compresi conseguimenti nell'ambito dell'elaborazione delle immagini da parte di un ampio sottordine di minuscoli pipistrelli. | << | < | > | >> |Pagina 249I fisici riconoscono quattro forze fondamentali della natura, due delle quali, la gravità e la forza elettromagnetica (EM), sono importanti per i biologi e gli ingegneri, mentre le rimanenti restano quasi sempre imprigionate all'interno dei nuclei atomici. La forza elettromagnetica si manifesta sotto tre forme: come campo elettrico, come campo magnetico, o come una combinazione dei due nota come radiazione EM. Nel loro insieme, gli appartenenti al regno animale sono sensibili a tutte e tre le manifestazioni, e le usano per esplorare il mondo all'intorno. In questa sede ci interessiamo principalmente di ricevitori EM passivi (gli «occhi»), perché è attraverso di essi che più comunemente gli animali sfruttano il campo EM. Talune creature si servono del campo elettrico, e le passiamo brevemente in rassegna alla fine del capitolo; altre possiedono un senso magnetico, e nel capitolo 11 vedremo come alcuni animali migratori percepiscono il campo magnetico terrestre. La luce è radiazione EM. E la luce visibile è quella piccola parte dello spettro elettromagnetico alla quale, per motivi che si chiariranno tra poco, i nostri occhi sono sensibili. Prima però di parlare degli occhi dobbiamo fare un ripasso di ottica, che è la fisica della luce. Al confronto dell'acustica del capitolo 8, è però una fisica molto meno complessa: le lunghezze d'onda coinvolte sono talmente corte che, salvo per una o due rilevanti eccezioni, possiamo dimenticare che la luce sia un fenomeno ondulatorio, e trattarla semplicemente come radiazione che segue percorsi rettilinei. Questo approccio, che è quello dell' ottica geometrica, è del tutto standard e universalmente accettato, a patto di ricordare che questa semplificazione si applica soltanto perché le lunghezze d'onda luminose sono di gran lunga inferiori alla scala di lunghezza di qualunque oggetto con il quale interagiscono, come le lenti o la retina. La figura 77 introduce ai termini e ai fenomeni ottici (la maggior parte) che dobbiamo conoscere. Le proprietà ottiche di un materiale trasparente sono descritte dal suo indice di rifrazione. Quando la luce passa da un mezzo trasparente a un altro con diverso indice di rifrazione, viene deviata in maniera predicibile. Se la transizione è brusca, come avviene all'interfaccia di una lente, il fenomeno non è graduale ma la traiettoria devia bruscamente, e da una retta in una direzione si passa a una retta in direzione diversa. (Le lenti biologiche hanno un indice di rifrazione prossimo a quello dell'acqua, n = 1,33, mentre quello dell'aria è n =1,00). La figura 77a mostra in che modo l'azione rifrangente produce la messa a fuoco di un'immagine. I raggi luminosi paralleli all'asse ottico vengono focalizzati dalla lente su un piano focale. Quelli emessi da oggetti molto distanti, quando raggiungono il sistema, in questo caso l'occhio, sono praticamente paralleli (linee continue), e quindi le immagini di questi oggetti si formano in sostanza sul piano focale; oggetti più vicini sono messi a fuoco oltre tale piano (linee tratteggiate). La forma di alcune lenti biologiche è modificabile (questa capacità è particolarmente sviluppata nei primati e nei carnivori, meno in altri ordini) , il che mette in gioco ulteriori elementi di ottica. La fisica della rifrazione ci dice che l'entità della deviazione non dipende solo dagli indici di rifrazione della lente e del mezzo, ma anche dall'angolo con cui la luce incide sulla lente o ne esce. Variando la distanza dell'oggetto varia l'angolo sotto cui i raggi entrano nella lente e quindi varia la distanza alla quale l'immagine viene focalizzata, come mostra la figura 77a. È per questo che noi vediamo a fuoco di volta in volta solo a una data distanza: i muscoli oculari automaticamente regolano la curvatura della lente, come mostra la figura 77b, in modo che i raggi emessi da oggetti posti a quella distanza convergano sulla retina, dove si trovano i nostri fotorecettori. Una lente perfetta focalizza la luce incidente in un solo punto, mentre se è difettosa può succedere che raggi che giacciono su piani diversi vengano messi a fuoco in punti diversi. Per esempio, la figura 77b mostra raggi nel piano del foglio convergenti in un certo fuoco; se due raggi in un altro piano convergono altrove, chiamiamo quella lente astigmatica.
Le cause di una messa a fuoco imperfetta, anche in assenza di difetti
della lente, sono essenzialmente due. La lente più semplice da costruire, si
tratti dell'occhio o dell'obiettivo della macchina fotografica, è
quella sferica, e in figura 77c potete vedere che lenti di questo tipo non
mettono a fuoco tutti i raggi nello stesso punto: il piano focale varia infatti
con la distanza dei raggi dall'asse ottico. Questo fenomeno, che
incontreremo ancora, è detto
aberrazione sferica.
La figura 77d illustra un altro effetto importante:
l'aberrazione cromatica;
questa è dovuta al fatto che l'indice di rifrazione varia con la lunghezza
d'onda della luce, cosicché una data lente ha lunghezze focali leggermente
diverse per luce di differenti colori. Le piccole lunghezze d'onda vengono
rifratte in misura maggiore; con luce blu la lunghezza focale è minore che con
luce rossa.
Ci interessano occhi in grado di formare immagini, non solo cellule sensibili alla luce, e questa restrizione ci porta a considerare tre soli phyla animali: i cordati, gli artropodi e i molluschi. Indaghiamo qui gli aspetti ingegneristici degli occhi animali, più che la loro struttura biologica. Per noi gli occhi sono sistemi di lenti che raccolgono informazioni su quanto sta intorno a un animale, per il tramite della radiazione EM dell'ambiente. Un occhio semplice ha un 'unica lente. Alcuni occhi di questo tipo (l'occhio umano e quelli della maggioranza dei vertebrati e di alcuni molluschi) mettono a fuoco l'immagine cambiando la forma della lente; altri (gli occhi di pesci, anfibi, serpenti e cefalopodi) spostano la lente, in modo simile alle macchine fotografiche. Alcuni animali hanno più di una lente in ciascun occhio e possono vedere immagini ingrandite; i loro occhi assomigliano a teleobiettivi. Incontreremo più avanti una di queste creature. La maggior parte degli animali sono artropodi che possiedono occhi composti formati da migliaia di lenticole. Per certi versi occhi di questo tipo sono meno efficienti degli occhi semplici, ma come vedremo hanno anch'essi i loro punti di forza. Per molti animali gli occhi rappresentano «il primario avamposto sensoriale del cervello». Si sono evoluti molte volte indipendentemente e sono bene adattati ad abitudini di vita e condizioni di illuminazione molto diverse (la varietà di forme di occhio che ne risulta è resa evidente in fig. 78) . Esistono per esempio almeno otto tipi eterogenei di occhi che si sono evoluti in maniera autonoma (quelli con lente a indice di rifrazione variabile che diminuisce con la distanza dall'asse ottico per correggere l'aberrazione sferica). Esistono undici modi conosciuti di combinare le lenti nell'occhio per produrre immagini a fuoco, e sei sono stati scoperti solamente nell'ultimo venticinquennio. La vista, com'è evidente, è un ausilio alla sopravvivenza di grande utilità, e la spinta evolutiva a svilupparne e migliorarne gli organi è molto forte. Esaminiamo quindi la diversità e i punti di forza dei vari schemi costruttivi di occhio, e ciò che vi sta dietro.
Gli occhi dei vertebrati si sono evoluti una sola volta, e quasi certamente in mare. I biologi produrranno tutti i convincenti riscontri del caso, ma per un ingegnere l'origine pelagica è un'ovvia conseguenza delle proprietà ottiche dell'acqua marina. Riportiamo in figura 79 l'attenuazione che subisce la radiazione EM nel propagarsi in quel mezzo; ricordando la definizione del decibel (dB) data nel capitolo 8, potete constatare ciò che la figura significa: la radiazione EM di quasi ogni frequenza viene completamente arrestata in pochi centimetri d'acqua, e solamente la stretta banda della luce visibile penetra oltre (per qualche decina di metri). Anche l'aria attenua la radiazione EM, ma in misura incomparabilmente minore. Il fatto che gli occhi dei vertebrati siano sensibili alla ristretta gamma di lunghezze d'onda che penetra nell'acqua marina suggerisce un'origine oceanica per il nostro senso della vista. La geometria di base dell'occhio semplice è illustrata nella figura 80. La luce penetra nel bulbo oculare attraversando la cornea trasparente e successivamente il cristallino, fino alla retina posta al fondo del bulbo. La cornea degli animali terrestri rifrange la luce ed è quindi di ausilio al cristallino nel mettere a fuoco le immagini. Nella maggior parte degli animali la cornea ha forma fissa e rifrange sempre nella stessa misura, ma le aquile e altri uccelli rapaci diurni ne possono variare il profilo così come fanno con il cristallino, in modo da regolare con precisione la messa a fuoco sulla retina. Sempre in figura 80 a e 80b vediamo come il cristallino cambi la curvatura (accomodazione) per mettere a fuoco sulla retina i raggi di luce incidenti. La figura 80c mostra inoltre una particolarità fondamentale dell'occhio a lente semplice: l'immagine che si forma sulla retina, anche se perfettamente a fuoco, è capovolta (e ribaltata tra destra e sinistra) una conseguenza scomoda ma inevitabile di questo tipo di occhio. Fortunatamente il cervello inverte l'immagine una seconda volta, in modo da mostrarci il mondo esterno nell'orientamento giusto. C'è molto altro da dire sulla considerevole mole di postelaborazione che ha luogo dietro gli occhi, ma limitiamoci per ora alle nozioni base dell'ottica. Quanto bene vedono occhi semplici? Ci sono svariate misure, la cui importanza biologica tuttavia varia da specie a specie: Acutezza (capacità di messa a fuoco). - Sensibilità (capacità di funzionare con diverse intensità luminose; un occhio sensibile vede con poca luce). - Discriminazione cromatica (capacità di distinguere le componenti spettrali della luce). Campo visivo (estensione dell'area abbracciata dall'occhio in ogni momento).
Percezione della profondità (capacità dell'occhio di fornire
informazioni sulla distanza di un oggetto).
|