|
|
|
|
|
|
|
| << | < | > | >> |Indice
Prologo 9
I. Il pianeta delle scimmie e l'antenato 15
- Il mondo dei Primati.
- Intorno a 50 milioni di anni fa.
- Le rivoluzioni scientifiche di Thomas Kuhn.
- Il pitecantropo e il caso Piltdown.
- L'autore della frode di Piltdown.
- Crollo di un paradigma.
- L'anello mancante non esiste.
- La forza debole dell'antropologia.
II. Australopithecus & C. 37
- Lucy in the Sky with Diamonds.
- Variabilità di una specie estinta.
- Sulle tracce dell'«harem» di Lucy.
- L'osso che non abbiamo.
- Lo scheletro nella roccia.
- Ancora un australopiteco?
- Tre minuscole ossa antichissime.
- L'ultimo degli Australopithecus?
- Australopithecus sediba, forse Homo.
III. Alle origini di Homo 61
- Phillip Vallentine Tobias.
- Homo habilis, since 1964.
- La diversità dei primi Homo.
- Alle falde del Caucaso.
- I fossili di Dmanisi.
- Il piccolo popolo dell'isola di Flores.
- Una nuova stella fra i primi Homo?
- Morfologia di una specie arcaica.
- La (sorprendente) datazione di Homo naledi.
IV. Umani del tempo di mezzo 85
- Atapuerca, la Sima de los Huesos.
- I fossili della Gran Dolina.
- Ancora su Homo antecessor.
- Il più antico europeo.
- Homo heidelbergensis.
- Denisova, sui monti Altai.
- Una misteriosa umanità estinta.
- I crani mancanti di Denisova?
V. Siti e manufatti preistorici 109
- Oltre cent'anni di Olduvai.
- I tesori di Melka Kunture.
- Manufatti davvero molto antichi.
- Un'altra storia di impronte.
- Vicino al villaggio di Buia.
- Il mistero dell'Acheuleano.
- Le penne dei Neanderthal di Fumane.
VI. Orologi e molecole 129
- Un orologio molecolare più lento.
- Regolando ancora le lancette.
- Analisi del DNA antico.
- «Scappatelle» coi Neanderthal.
- Quando ci siamo incrociati?
- Epigenetica dell'antenato.
VII. Neanderthal 147
- Glaciazioni e crisi demografiche.
- I dentini della baia di Uluzzo.
- Quelle «scappatelle» levantine.
- L'hanno fatto anche in Europa?
- Dentro la zona ibrida.
- Speleologi e misteriosi architetti.
- Neanderthal cannibali.
- Glaciazioni e aree rifugio.
VIII. Storie italiane 171
- Nuova data per il cranio di Ceprano.
- A Rebibbia c'è «un mammut».
- L'uomo di Altamura.
- Nuove ricerche ad Altamura.
- Il volto dell'uomo di Altamura.
- Che cosa racconta grotta Guattari.
- Breve storia dell'uomo venuto dal ghiaccio.
- Il cranio che ispirò Cesare Lombroso.
IX. Homo sapiens 193
- Gould e le origini di Homo sapiens.
- L'unicità del nostro volto.
- Lo sviluppo encefalico di Homo sapiens.
- Un cervello pieno di odori.
- Storie di denti e cervello.
- La prima domesticazione: il cane.
- I più antichi americani.
- «Ritorno» in Africa nel Neolitico.
- Le razze umane non esistono.
Epilogo 219
Figure 225
Bibliografia 231
|
| << | < | > | >> |Pagina 9Dove si dichiara di voler fare in modo che il passaggio finale delle ricerche sull'evoluzione umana sia la divulgazione; così da condividere con tutti, ma proprio tutti, una storia che è di tutti. Qui si parla del nostro passato «remoto»: quello degli ominidi bipedi e pelosi, quello di Lucy e dei Neanderthal, quello delle grandi diffusioni dall'Africa verso l'Eurasia, quello dei crani fossili, degli scheletri frammentari e dei siti preistorici, quello dei manufatti del Paleolitico, quello della paleogenetica. Conoscere la nostra storia anche la più antica, di quando non eravamo nemmeno umani è una consapevolezza che non guasta, soprattutto oggi che siamo i padroni (incontrollati) del pianeta; perché dentro di noi c'è sempre quel bipede barcollante che, intorno a 2 milioni di anni fa, iniziò a sviluppare un cervello abnorme, e poi, circa 200 mila anni or sono, divenne Homo sapiens e si diffuse ovunque. Divulgare, o «disseminare» (come dicono gli inglesi). Questo è il senso profondo, a me pare, del lavoro che faccio. Noi paleoantropologi passiamo settimane immersi nella terra, con in mano pale, pennelli, bisturi e setacci, poi emergiamo dai quadrati dei nostri scavi per ricomparire in laboratori che ormai assomigliano a quelli della polizia scientifica, attrezzati con complessi apparecchi per l'amplificazione e l'analisi di DNA degradato oppure con scanner laser e tomografi per la digitalizzazione di immagini tridimensionali. Infine passiamo gran parte del nostro tempo davanti a computer sempre più miniaturizzati per analizzare dati, elaborare immagini, raccontare le storie che siamo riusciti a ricomporre da evidenze frammentarie che lo stesso Darwin, con una metafora efficace, paragonava a pezzetti di pagine strappate di un libro da restaurare. Che senso avrebbe tutto ciò, se non ci fosse un ultimo passaggio, quello della divulgazione? Da qui l'imperativo: raccontare, raccontare, raccontare... E non è certo una storia da tenere all'interno di una comunità scientifica che, su scala mondiale, non supera qualche migliaio di persone. Il significato vero del nostro lavoro è che queste storie vanno raccontate a tutti; sì, proprio a tutti. È qui che l'antropologia (nel suo senso più ampio) assume una valenza culturale, sociale e, se vogliamo anche politica. Qualche tempo fa, ad esempio, mi è capitato di sedermi a una tavola rotonda organizzata da un prestigioso ateneo romano, insieme ad alcuni illustri colleghi (un etologo, un filosofo e due giuristi), per commentare un poderoso trattato di Sergio Ortino dal titolo La struttura delle rivoluzioni economiche (2010). Quando il libro era arrivato nel mio studio all'università avevo pensato a un errore postale. Poi, una volta aperto e visti i contenuti, ho capito. I primi capitoli la metà almeno di un volume di 700 pagine sono zeppi di citazioni della nostra scienza, la scienza delle nostre origini, traboccano di evoluzione umana. L'autore è convinto infatti (e io con lui) che la nostra storia evolutiva sia essenziale per comprendere le vicende storiche del passato più recente dell'umanità, con le varie rivoluzioni economiche e culturali che l'hanno attraversato, ma anche il nostro presente, che è già futuro: il futuro dell'informazione globale e delle sfide del mondo contemporaneo. Come dargli torto? Gli aggiornamenti in pillole che troverete nelle prossime pagine una sorta di naturale continuazione di altri libri e, in particolare, del mio Il grande racconto dell'evoluzione umana , queste brevi «ultime notizie» saranno anche, di volta in volta, l'occasione per fare un po' di luce sui rami dell'albero frondoso della nostra storia naturale e per provare a capire qualcosa di più riguardo ai tanti argomenti su cui la paleoantropologia ha galoppato negli ultimi anni. Con questo taglio narrativo, vorrei farmi guidare da un impareggiabile maestro (senza la minima presunzione di eguagliarlo): Stephen Jay Gould. Non credo che ci sia bisogno di ricordare ai miei lettori chi è stato Gould, scomparso prematuramente il 20 maggio 2002. Basterà dire che stiamo parlando di uno dei più importanti paleontologi e biologi evoluzionisti della seconda metà del XX secolo. Da paleontologo, insieme a Niles Eldredge anche lui paleontologo, all'American Museum of Natural History di New York ha contribuito in modo decisivo a riconsiderare alcuni meccanismi dell'evoluzione darwiniana, proponendo la teoria dei punctuated equilibria, cioè «equilibri intermittenti» o «puntuati» o «punteggiati». Con Elisabeth Vrba altra paleontologa, alla Yale University ha introdotto il termine exaptation, una rivisitazione del concetto già introdotto da Darwin come «pre-adattamento». Il suo libro del 1977, Ontogenesi e filogenesi (Ontogeny and Phylogeny), ha dato un impulso fondamentale allo sviluppo dell'approccio cosiddetto evo-devo, in base al quale si combinano le conoscenze su accrescimento e sviluppo (ontogeny, appunto) con quelle sull'evoluzione (phylogeny). Infine, con il monumentale trattato La struttura della teoria dell'evoluzione vero e proprio suo testamento scientifico ci ha lasciato una delle opere fondamentali sulla teoria dell'evoluzione dopo L'origine delle specie di Charles Darwin. Per quanto sia di mia conoscenza, Gould non si impegnò mai attivamente nel campo della ricerca antropologica o paleoantropologica, tranne che in un articolo del 1974 (scritto a due mani con David Pilbeam e pubblicato su «Science»). Tuttavia la sua influenza sullo studio della nostra specie e sulla nostra visione delle origini degli ominidi e dell'evoluzione umana in genere è stato straordinario. Non voglio dilungarmi, ma mi pare evidente come quanto ho appena brevemente elencato possa aver avuto un impatto formidabile su quello che oggi pensiamo della nostra storia naturale, come pure su alcune linee recenti della ricerca. Tanto per fare un esempio, non vediamo più l'evoluzione umana come una sequenza lineare di specie che si sono succedute nel tempo una dopo l'altra, ma come una storia complessa e ramificata, cioè come un «albero frondoso» per usare un'espressione dello stesso Gould, o un «cespuglio» come dicono molti. Non finisce qui. Gould ha anche svolto magistralmente il ruolo di divulgatore scientifico. Coniugando arguzia e leggerezza con sapienza e rigorosa capacità di sintesi, ha reso memorabili i suoi celebri interventi mensili su «Natural History», successivamente raccolti in volumi tradotti in molte lingue e divenuti anche da noi molto popolari. Da questi scritti traspare, fra l'altro, tutta la sua avversione verso la mistificazione, di ogni tipo e natura: contro la pseudoscienza a sostegno del razzismo o l'insegnamento del creazionismo nelle scuole statunitensi trattato come se fosse una teoria scientifica. Ritroverete un po' dell'eredità di Gould nelle prossime pagine...
| << | < | > | >> |Pagina 23Le rivoluzioni scientifiche di Thomas KuhnPiù di cinquant'anni fa, nell' Encyclopedia of Unified Science della Chicago University Press si pubblicava un volumetto che, sulla carta, non era destinato ad avere un particolare successo. E così all'inizio effettivamente andò. Tuttavia col tempo le vendite si impennarono, e quel saggio in origine rivolto ai cultori della materia è diventato un long seller da oltre un milione di copie. Non solo. Da quelle pagine è scaturito un lessico che si è diffuso ben oltre i confini della filosofia della scienza, dal momento in cui tutti (o comunque in tanti) ci siamo spesso riempiti la bocca con termini come «paradigma», «scienza normale», «rivoluzione scientifica» e così via. L'autore del saggio era Thomas Kuhn , classe 1922 e scomparso nel 1996, fisico di formazione e storico della scienza per professione. Il libro aveva effettivamente un titolo da pietra miliare La struttura delle rivoluzioni scientifiche ( 1962) e tale è diventato davvero, mettendosi in competizione con l'empirismo logico del Circolo di Vienna o con il razionalismo critico di Karl R. Popper. La proposta era a prima vista semplice. Secondo Kuhn la storia mostra che il progresso scientifico non procede come una marcia verso la verità, ma piuttosto avanza a salti che possiamo chiamare (parole sue) «slittamenti di paradigma», dove «paradigma» è un sistema coerente di visioni della realtà che, in un certo momento storico, è condiviso dalla comunità scientifica di riferimento e tale rimane fino al paradigma successivo. Dunque, per fare l'esempio forse più banale, dopo Tolomeo il paradigma è stato che la Terra sia al centro dell'Universo, mentre dai tempi di Copernico è stato il Sole a prenderne il posto. Nel tempo la ricerca si muove per lo più su un terreno che potremmo dire pianeggiante, dove si indagano le conseguenze del paradigma condiviso, in una fase di relativa stasi denominata da Kuhn «scienza normale»... Fino a quando i conti non tornano più e si attraversa una «fase rivoluzionaria», conclusa dall'affermazione di un nuovo paradigma che a sua volta precede la successiva fase di scienza normale. In questo modo viene introdotta nella storia e nella filosofia delle scienze, anche delle cosiddette scienze più «dure», una quota di relativismo storico-culturale. Come dire che la scienza, lungi dall'essere asettica, è invece profondamente condizionata dal clima culturale della sua epoca e dalla circolazione di idee a cui essa stessa partecipa. Questo taglio storicista non è piaciuto a molti, e credo che sia anche per questa ragione che il contributo epistemologico di Thomas Kuhn è stato criticato da più parti e ancora oggi rimane controverso. Tranquilli. Non è nelle mie corde (né è il mio mestiere) entrare in questo dibattito, e dunque mi fermo qui. Vorrei piuttosto valutare insieme a voi se e in quale misura i contenuti di fondo della proposta di Kuhn possano trovare un'applicazione nel campo della paleoantropologia. Ovvero se la storia della ricerca sulle nostre origini possa essere letta in termini di rivoluzioni, paradigmi e fasi di scienza normale. Non è questa la sede per affrontare in maniera esaustiva una questione di tale rilevanza. Dobbiamo però parlarne in modo più approfondito, per vedere come l'idea ottocentesca di «anello mancante» possa aver portato all'aberrazione della celebre frode di Piltdown, presto smentita dalle prove che si andavano accumulando in Africa a partire dalla scoperta del primo Australopithecus (tutt'altro che un essere umano dal grande cervello con la faccia da scimmia) o come, per mezzo secolo, il paradigma della teoria sintetica dell'evoluzione abbia guidato i ricercatori a credere nell'esistenza (erronea, pensiamo oggi) di una sola specie umana che, gradualmente, avrebbe evoluto caratteristiche moderne a partire da quelle arcaiche.
A presto, dunque: basta voltare pagina.
Il pitecantropo e il caso Piltdown
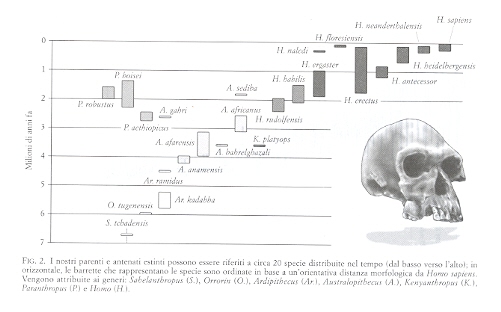
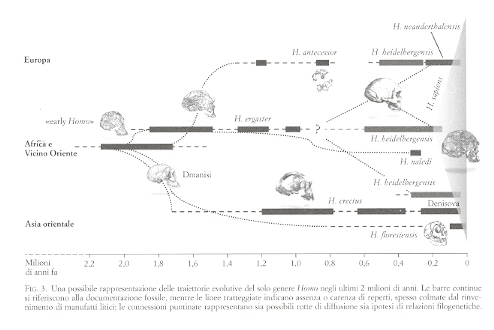
Dicevamo di Thomas Kuhn e del suo libro del 1962 La struttura delle rivoluzioni scientifiche, che tanta parte ha avuto nella discussione epistemologica degli ultimi cinquant'anni. Abbiamo poi espresso l'intenzione di applicare il modello di Kuhn basato su un'alternanza fra salti «slittamenti» di paradigma e fasi di «scienza normale» ad almeno un paio di esempi tratti dalla storia della paleoantropologia. Vediamo il primo esempio. Nella seconda metà dell'800 poteva essere appropriato parlare di anello mancante, quando i resti fossili rinvenuti non erano altro che alcuni Neanderthal e Cro-Magnon; troppo recenti ed encefalizzati per poter rappresentare l'antenato. Pertanto, affrontare fino in fondo il tema del nostro posto nella natura era possibile solo sulla base delle affinità fra noi e le scimmie antropomorfe. Così facendo mancava però un anello, almeno uno, alla catena dei viventi. Ecco l'idea: un anello mancante. | << | < | > | >> |Pagina 29Crollo di un paradigmaUn altro interessante esempio dell'alternanza fra «rivoluzioni scientifiche» e fasi di «scienza normale» ha pervaso la ricerca delle nostre origini per buona parte del '900: si tratta del cosiddetto «paradigma della specie unica». E l'uso del termine «paradigma» è già un esplicito riferimento al modello epistemologico proposto da Thomas Kuhn nel 1962. Agli inizi del secolo scorso i paleoantropologi avevano preso un brutto vizio: davano un nome in latino a ogni fossile che trovavano. La denominazione linneana di genere e specie perdeva così di significato e di valenza interpretativa. Era solo un'etichetta da mettere sotto il reperto nella vetrina di un museo. Poi, intorno agli anni '30, quello che Julian Huxley definì «oblio del darwinismo» si concluse con la saldatura tra la selezione naturale di Darwin e la genetica nata dalla riscoperta delle leggi di Mendel , decretando l'esordio della moderna teoria sintetica dell'evoluzione. In questa fase, che potremmo dire infantile, si era portati però a confondere un po' troppo il fenomeno intraspecifico della microevoluzione con quello sovraspecifico della macroevoluzione. La genetica di popolazioni era il deus ex machina, con le sue variazioni di frequenze alleliche, le curve a campana, le diverse pressioni selettive stabilizzanti e direzionali. Si pensava che un simile cambiamento graduale si applicasse bene anche alla macroevoluzione, come se questa soggiacesse agli stessi meccanismi, come se non fosse altro che una microevoluzione sviluppata su tempi grandiosi. E i paleoantropologi, strigliati a dovere da uno dei padri della teoria sintetica ( Ernst Mayr ), si adeguarono. La pletora di nomi in latino dei primi decenni del XX secolo sparì e si passò a una più sobria distinzione in due soli generi (Australopithecus e Homo), con poche specie al loro interno. Si affermò l'idea di un'evoluzione umana lineare e sequenziale, ben rappresentata da quella sfilata di scimmie, scimmioni e umani che ancora vediamo sui giornali ogni volta che si parla delle nostre origini. Stephen J. Gould le chiamava «iconografia della speranza»: una sorta di marcia verso il progresso, dove una specie segue l'altra e ciascuna occupa da sola una certa fase storica, un preciso grado dell'evoluzione umana. Si trattava, appunto, del paradigma della specie unica. E il finalismo, dietro le quinte, gongolava, visto che si legittimava l'uso di termini come «ominazione» o (peggio) «sapientizzazione», che chiaramente alludono a una visione teleologica. Chi interpretava l'evoluzione umana non poteva non tenere conto di questo paradigma. Nacquero così il modello detto «a candelabro» e la sua versione recente, detta «evoluzione multíregionale». In entrambi i casi, pur di avere una sola specie alla volta come dire una specie unica in costante, graduale evoluzione, lanciata verso la modernità si doveva ammettere un'origine poligenica della variabilità umana attuale o un'improbabile gittata transcontinentale del flusso genico. Tutto a posto? Non proprio. Dopo una serie di scosse telluriche dovute all'intervento di nuove idee, di nuovi fossili e di nuove prove sperimentali, a cavallo degli anni '80 del '900 il paradigma è caduto. Adesso il modello è un altro e vediamo l'evoluzione umana non più come una sequenza lineare, ma come un percorso complesso e ramificato. Si parla allora di «cespuglio» (anche se a me pare più corretta la metafora dell'«albero frondoso» che lascia meno spazio a suggestioni polifiletiche) e i paleoantropologi sono tornati a contare molti generi, con almeno una ventina di specie. Non più per mettere cartellini nelle vetrine dei musei, ma per rendere conto di una variabilità preistorica sempre più sorprendente, che ci porta a raccontare una storia complessa. Siamo dunque ormai in una nuova fase di scienza normale? | << | < | > | >> |Pagina 34La forza debole dell'antropologiaSe provo a fare un bilancio di ciò che scrivo in queste pagine o di quello che racconto nelle conferenze che mi capita di tenere in giro per l'Italia, mi accorgo che, pur non essendoci una precisa intenzione da parte mia, mi trovo spesso a suggerire cautela verso facili entusiasmi o a mostrare che, con il passare del tempo, certe ipotesi possono essere riconsiderate o, magari, abbandonate. D'altra parte, funziona proprio così. In scienze come quella delle nostre origini si procede per modelli, teorie e ipotesi di lavoro... La ricerca non porta a soluzioni definitive, né tanto meno a leggi, mentre le ipotesi, in quanto tali, dovranno essere verificate da nuove ricerche. Elaborare su di esse speculazioni a briglia sciolta può essere dunque prematuro e fuorviante. Con la scienza delle nostre origini (o paleoantropologia) tutto ciò accade ancora di più. E non solo perché essa si basa su fragili prove, rappresentate da ossa frammentarie, denti spesso isolati e manufatti di pietra dispersi nello spazio e nel tempo. Il problema lo si ritrova nel più vasto campo disciplinare dell'antropologia (cosiddetta «fisica»). È questo l'interesse, il fascino e la forza, ma anche la debolezza e, direi, la condanna delle scienze dell'uomo. E già, visto che si tratta di noi stessi, tutti siamo interessati alla nostra storia e alla nostra natura (da qui il fascino e la forza), ma ci sentiamo anche ispirati a proporre conclusioni talvolta affrettate (da qui la debolezza e la condanna), inevitabilmente agganciate alla nostra formazione e alle nostre personali esperienze. Al lavoro dei ricercatori e ai loro modelli si aggiunge, allora, un rumoroso coro di voci incontrollate e incontrollabili, che esprimono il punto di vista più che legittimo, ma quasi sempre tutt'altro che scientifico di operatori di altri settori del sapere e della comunicazione. | << | < | > | >> |Pagina 61Dove si scopre che, ancora in Africa, intorno a 2,5 milioni di anni fa e in virtù degli adattamenti acquisiti in precedenza, la storia cambiò passo e si andarono affermando e diffondendo diverse varietà di primi rappresentanti del genere «Homo». Per non cadere in equivoci, o addirittura in errore, ci siamo abituati a chiamarli provvisoriamente «early Homo». In effetti, i più antichi fossili ascrivibili al genere Homo altro non sono che rappresentanti della più antica e arcaica specie umana (cioè Homo habilis) e/o di forme affini. Ma il quadro è assai più complesso e problematico di quanto possa sembrare a prima vista. Vediamo meglio. Dopo la denominazione di Homo habilis, nel 1964, i resti fossili e i manufatti riferibili a questa prima specie umana sono molto aumentati di numero e di qualità. Così, è necessariamente anche aumentata la complessità dello scenario evolutivo che si è presentato via via davanti ai nostri occhi. Data l'importanza di questa fase dell'evoluzione umana cruciale snodo fra il mondo di Australopithecus e la nuova biologia ed ecologia del genere Homo ci sono state perciò e sono tuttora in corso diverse revisioni dell'evidenza fossile disponibile e di tutte le altre informazioni che abbiamo. Ma c'è dell'altro. Fra gli «early Homo» dobbiamo probabilmente inserire anche la più recente specie di Australopithecus (di cui abbiamo parlato), Australopithecus sediba, oppure l'ultima star fra gli ominidi bipedi nostri parenti estinti, ovvero l'ancora enigmatico Homo naledi, rinvenuto anch'esso (e con un gran numero di reperti fossili) in un'ennesima grotta carsica del Sudafrica.
Tuttavia, al momento, malgrado la quantità delle evidenze disponibili anche in termini dei primi manufatti del Paleolitico, il risultato è che sull'origine, sulla collocazione tassonomica e sull'esatta posizione filogenetica dei primi esseri che possano dirsi «umani» non esiste ancora né piena luce né tanto meno concordia fra gli specialisti. È per questo che al momento ci semplifichiamo la vita usando l'espressione sintetica «early Homo» piuttosto che ricorrere a un guazzabuglio di binomi in latino, della cui effettiva validità non siamo nemmeno tanto sicuri. Ciò di cui siamo abbastanza sicuri è che con la comparsa degli «early Homo» si impostano un nuovo modello biologico e una nuova collocazione ecologica dei nostri antenati. Questi mostrano di conservare uno scheletro non dissimile da quello dei loro contemporanei Australopithecus e Paranthropus, mentre rilevanti cambiamenti sono intervenuti a carico della morfologia cranica e della dentatura. Osserviamo infatti un primo aumento delle dimensioni encefaliche (con un volume superiore, anche se di poco, a mezzo litro), associato a una struttura dentaria non così specializzata come quella delle australopitecine. A questi cambiamenti si affiancano un nuovo assetto della volta cranica, una riduzione delle sovrastrutture ossee, con ossa facciali e mandibola più gracili. Ma soprattutto, avendo cambiato posizione nella piramide alimentare da consumatori di una varietà di alimenti vegetali, come furono le australopitecine, a una dieta con maggiore apporto di proteine animali, cioè carnivora i nostri antenati acquisiscono il ruolo di predatori opportunisti (si parla di «spazzini della savana»), quasi questo fosse un preludio delle attitudini da cacciatori che altri Homo diverranno poi. In rapporto a questa nuova nicchia ecologica, gli «early Homo» avevano anche raggiunto, ben prima di 2 milioni di anni fa, una piena abilità manuale e la capacità di produrre manufatti: quelli del cosiddetto «Modo 1» del Paleolitico, o Olduvaiano (che prende nome dal ben noto sito di Olduvai in Tanzania). È probabile perciò che, a partire da questa cruciale fase di passaggio della nostra evoluzione, due fra le più importanti caratteristiche che distinguono l'uomo dagli altri Primati si siano incrementate a vicenda: il fenomeno noto come encefalizzazione, costituito dal progressivo aumento delle dimensioni encefaliche, con espansione della scatola cranica, e la produzione di strumenti in pietra archeologicamente ben riconoscibili, che ha proiettato questi nostri antenati nella dimensione del Paleolitico. Inoltre, è proprio da questo momento, cioè da circa 2 milioni di anni fa, che la nostra storia cessa di essere una vicenda che ha come scenario esclusivo l'Africa, anzi le regioni orientali e meridionali dell'Africa subsahariana. Tra breve non parleremo più solo di questa, ma anche di altre aree africane più a nord e di una varietà di territori in larga parte dell'Eurasia, che a questo punto si rendono disponibili e diventano lo sterminato scenario geografico della nostra evoluzione. Così, la prima e altre successive diffusioni degli ominidi vengono chiamate out-of-Africa, visto che spesso avvennero in modo centrifugo da quel continente verso l'Eurasia. C'è chi le ha numerate queste diffusioni e le principali sarebbero due: out-of-Africa 1 e out-of-Africa 2 (quella cioè, di molto successiva, dei primi rappresentanti della nostra specie). Personalmente preferisco evitare questa semplificazione, in quanto ho l'impressione (più che un'impressione, come vedremo) che di diffusioni out-of-Africa ce ne siano state ben più di due. Anzi, l'intera storia del genere Homo, nel corso di almeno 2 milioni di anni è stata una successione quasi continua di diffusioni geografiche. La prima grande diffusione, prima in Asia e poi anche in Europa, comporterà la formazione di una varietà di ominidi bipedi dal cervello sempre più grande, che saranno ormai tutte appartenenti allo stesso genere Homo. Anche il resto della storia sarà, come la precedente, tutt'altro che lineare e scopriremo presto le numerose «variazioni sul tema» che hanno punteggiato l'evoluzione del nostro genere nel corso degli ultimi 2 milioni di anni, tanto in Africa quanto in Eurasia. Incontreremo così il prezioso campione fossile di Dmanisi, nel Piccolo Caucaso, ossia nel territorio dell'attuale Georgia, quasi fosse una tappa della diffusione dei primi Homo in Eurasia. Allo stesso tempo, vedremo evolversi in Africa i rappresentanti di una specie ormai inequivocabilmente «nostra», cioè Homo ergaster, mentre vedremo caratterizzarsi in Estremo Oriente la specie forse più nota fra gli umani arcaici, cioè Homo erectus, ma anche quella che rappresenta il piccolo e longevo popolo dell'isola di Flores, cioè Homo floresiensis, emerso a partire da creature simili agli «early Homo» in diffusione, che seguirà un suo peculiare percorso evolutivo, dopo essersi trovato in isolamento oltre quella formidabile barriera geografica che, per molte forme di vita e da tempo immemore, è stata la cosiddetta «linea di Wallace». | << | < | > | >> |Pagina 72Dmanisi si trova nel Piccolo Caucaso, in Georgia, a metà strada fra il mar Nero e il mar Caspio, a un'ora di macchina dalla capitale Tbilisi (circa 85 chilometri a sud-ovest). La posizione del sito è particolarissima: in cima a uno sperone roccioso che a sua volta sovrasta, come un gigantesco ferro da stiro, la confluenza fra due fiumi. In cima a questo sperone c'è una cittadella medievale sommersa dalla vegetazione, con tanto di ruderi del castello nella parte più alta e una deliziosa chiesetta ancora in funzione. Furono proprio gli archeologi che negli anni '80 stavano scavando fra le rovine medievali a trovare i denti di un rinoceronte. Un rinoceronte?! Abesalom Vekua, esperto paleontologo del Museo di Tbilisi, stabilì che quei denti non potevano certo essere di epoca medievale, ma che invece lì sotto dovevano esserci strati più antichi, parecchio più antichi. Iniziò così uno scavo paleontologico, a fianco di quello medievista. Presto venne alla luce una varietà di ossa, denti e corna di animali del primo Pleistocene. Insieme a queste faune dal sapore africano apparvero anche manufatti, testimoni della presenza dell'uomo, e nel 1991 una mandibola umana. All'inizio l'importanza della mandibola non fu capita in pieno, e il punto di vista del paleoantropologo georgiano Leo Gabunia il quale pensava si trattasse di una primordiale forma umana che, per prima, si era diffusa fuori del continente africano rimase inascoltato per quasi un decennio. Molti però gli diedero ragione quando, al cambio del millennio, da Dmanisi iniziarono come a «zampillare» fossili umani: cinque crani al momento e quattro mandibole, due omeri, due femori, una tibia, una scapola, tre clavicole e poi vertebre, costole, falangi e così via. Insomma, un campione fossile davvero eccezionale, vista anche l'antichità e le caratteristiche stesse dei reperti fossili. Il punto chiave per capire Dmanisi sta proprio nella combinazione fra geografia (al crocevia di tre continenti), datazione (una delle più antiche fuori dell'Africa), manufatti (decisamente primitivi) e morfologia dei fossili (che li avvicina agli Homo più arcaici). A quest'ultimo proposito i ricercatori georgiani, da un decennio sotto la guida di David Lordkipanidze, hanno proposto il nome di una nuova specie, Homo georgicus, sulla base del reperto più «stravagante» fra quelli rinvenuti: una mandibola indicata con la sigla D 2600.Chi era allora Homo georgicus (ammesso che questo nome abbia credito)? Probabilmente aveva ragione Leo Gabunia. Trattandosi di una primordiale forma umana che per prima uscì dall'Africa, porta con sé i caratteri dell'antenato (Homo habilis), ma al tempo stesso richiama quelli propri degli esseri umani che nel frattempo continuavano a evolvere in Africa (Homo ergaster) e annuncia quelli di specie che arriveranno poi sia in Estremo Oriente (Homo erectus) sia in Europa (Homo antecessor).
Un bel rebus, non c'è che dire.
I fossili di Dmanisi
Continuiamo a parlare del sito di Dmanisi, in Georgia: una finestra aperta su una delle fasi cruciali e più interessanti dell'evoluzione umana; alle origini del genere Homo, ma ormai fuori della culla africana; al crocicchio di tre continenti (Africa, Asia ed Europa) e con una datazione che si aggira intorno a 1 milione e 800 mila anni fa, cioè una delle più antiche fuori dell'Africa. Forse la più antica, o quanto meno la più solida. Vale senz'altro la pena di parlarne ancora, perché si tratta di uno dei siti migliori in tutto il panorama della paleoantropologia di ieri, di oggi e, con ogni probabilità, anche di domani. Anzi, l'importanza del sito, dei fossili e dei manufatti che vi vengono ritrovati è destinata a crescere, grazie agli studi sul materiale raccolto negli ultimi vent'anni e alle ricerche che sono tuttora attivamente in corso nel sito. E chissà che proprio mentre scrivo non stiano tornando alla luce nuovi reperti e altre importanti tracce preistoriche.
Quando si dice che i fossili umani di Dmanisi sono un rebus paleoantropologico dato che racchiudono in sé caratteristiche di diverse specie estinte del genere Homo non si sbaglia, né si esagera. Ma c'è di più. Dalle caratteristiche dei fossili di Dmanisi è principalmente emerso che i protagonisti della prima diffusione extra-africana del genere Homo non erano ominidi dal cervello voluminoso e dalle notevoli capacità tecnologiche o addirittura «culturali», visto che il volume endocranico dei fossili georgiani è decisamente modesto (siamo intorno a 700 millilitri) e considerata la tecnologia arcaica dei loro manufatti. Piuttosto, sembra sempre più probabile che a diffondersi fuori dell'Africa siano state forme primordiali del genere Homo, con elementi che richiamano ominidi ancora in parte scimmieschi, come Australopithecus. Su queste (e altre) basi si è inclini a ritenere che la prima diffusione del genere Homo abbia avuto una natura eminentemente ecologica (non culturale), dettata dai nuovi adattamenti di questi nostri remoti antenati, tra cui un incremento nel consumo di carne e l'uso di manufatti in pietra. Una traiettoria avrebbe così toccato le pendici meridionali del Caucaso, mentre altre si orientavano più a est verso l'Estremo Oriente. È probabile che barriere geografiche, fattori climatici e limiti biologici caratteristici di questi ominidi, originari delle savane africane abbiano influito sulla tendenza dell'onda di diffusione a muoversi dapprima lungo i paralleli, piuttosto che lungo i meridiani. Solo in seguito l'uomo arriverà anche in Europa. Un altro aspetto di grande interesse riguarda la diversità dei fossili umani rinvenuti a Dmanisi, con individui dei due sessi e di varia età. Quei crani, le mandibole e gli altri resti dello scheletro forniscono un raro quadro di variabilità, probabilmente interna a una stessa popolazione di primi Homo: qualcosa che sarebbe estremamente interessante trovare in Africa. L'area d'origine del nostro genere cioè l'Africa orientale, ma c'è chi guarda con attenzione anche al Sudafrica ha infatti restituito diversi siti con fossili riferibili alle prime specie del genere Homo, ma nessuno di essi fornisce un quadro di variabilità interna come quello di Dmanisi. Ciò comporta che, al momento, ci sia incertezza sul numero delle specie africane esistite fra 2,5 e 1,5 milioni di anni fa e su quali resti attribuire a ciascuna di esse. Anche per questo, le origini del genere Homo sono ancora avvolte, almeno in parte, dalla nebbia. Chissà che Dmanisi, come un «regolo» paleontologico, non ci aiuti a dissolvere un po' di quella nebbia africana... | << | < | > | >> |Pagina 98Homo heidelbergensisDa quando ci siamo addentrati nel sistema carsico della Sierra de Atapuerca, abbiamo scoperto tracce dell'evoluzione umana in Europa in fasi cruciali di questo percorso. Fra esse ci sono quelle di un primo, forse discontinuo ingresso nel continente da parte di forme umane arcaiche, a partire da almeno 1,2 milioni di anni fa. Ci sono poi quelle del più massiccio arrivo degli uomini dell'Acheuleano (il cosiddetto «Modo 2» del Paleolitico inferiore), a partire da circa 600 mila anni fa. Con questo secondo ingresso di popolazioni umane in Europa, entriamo in un altro territorio di frontiera della ricerca paleoantropologica contemporanea. Sì, proprio «di frontiera». Soltanto una ventina d'anni fa si trattava di un campo praticamente inesplorato o comunque poco noto, riferito a un'umanità vagamente ancestrale alla nostra, chiamata «Homo sapiens arcaico». Oggi abbiamo le idee decisamente più chiare (o, almeno, pensiamo di averle) sull'identità della nostra specie e sulle origini di Homo sapiens, che sono avvenute in Africa, 200 mila anni fa o giù di lì. Dunque, non ha più senso parlare di un fantomatico «Homo sapiens arcaico», ma semmai si deve fare riferimento a una specie differente, da considerarsi ancestrale rispetto a quella moderna. Per questa specie c'è un nome disponibile e questo nome è Homo heidelbergensis. Il binomio linneano venne coniato nel 1908, a seguito della descrizione di una mandibola fossile rinvenuta l'anno prima a Mauer, dalle parti di Heidelberg in Germania. Le caratteristiche di quella mandibola erano tali da non poter essere confuse con alcuna delle altre forme fossili di Homo allora conosciute in Europa: Neanderthal (Homo neanderthalensis) e Cro-Magnon (Homo sapiens). Dunque un nome nuovo: Homo heidelbergensis. La mandibola di Mauer ha circa mezzo milione di anni e, nel secolo trascorso dalla sua scoperta, molti altri fossili di analoga antichità sono stati rinvenuti in Europa, ma anche in Africa e Asia: mandibole, crani e altri resti dello scheletro. Nel loro insieme, sono reperti che condividono (oltre a una simile antichità) diversi tratti morfologici, sia pure in un quadro di variabilità interregionale favorita dal vasto areale. I caratteri in comune consentono, come nel caso della mandibola di Mauer, di escludere l'appartenenza a Homo sapiens, Homo neanderthalensis, Homo erectus (l'umanità arcaica dell'Estremo Oriente) o anche agli ultimi Homo ergaster africani, per non parlare dei primi europei (Homo antecessor) o del caso particolarissimo degli hobbit dell'isola di Flores in Indonesia (Homo floresiensis). Dunque il quadro sembra chiaro. C'è una sorta di «umanità di mezzo» (compresa fra almeno 600 e circa 200 mila anni fa), distribuita in Eurasia e in Africa, che si interpone tra forme umane più arcaiche (molte delle specie appena elencate) e varietà successive come i Neanderthal e noi stessi. Anzi, di più. Homo heidelbergensis è la specie ancestrale a entrambe queste ultime, l'antenato comune da tempo cercato. | << | < | > | >> |Pagina 129Dove si scopre che ci sono orologi le cui lancette sono fatte di molecole e sono regolati da distanze genetiche che mutano nel tempo; questo (e molto altro) lo possiamo ritrovare nel DNA dei vivi e in quello che si è conservato nei fossili. Le distanze genetiche cioè la misura della diversità genetica fra specie viventi, come ad esempio fra noi e gli scimpanzé negli ultimi decenni si sono rivelate estremamente utili nell'indicare i tempi delle principali ramificazioni dell'albero evolutivo. In base ad alcune assunzioni e a semplici algoritmi (di cui non staremo a dire), le distanze genetiche possono infatti tradursi in stime del tempo intercorso dall'inizio della separazione di due specie da uno stesso antenato comune. È un calcolo che ha preso il nome di «orologio molecolare», in quanto si basa su una valutazione delle divergenze filogenetiche a partire dal dato molecolare. Tempi ottenuti cioè da informazioni fatte di proteine, di DNA nucleare, di DNA mitocondriale (mtDNA) ecc. Molecole, insomma. Eravamo intorno al 1980 quando i paleoantropologi finalmente capirono cosa c'era dietro quelle stime dei tempi dell'evoluzione basate sulle molecole. Capirono e accettarono quanto i biologi molecolari impegnati a studiare i tempi dell'evoluzione dicevano dalla metà degli anni '60, affermando che le distanze genetiche fra specie attuali possono essere messe in rapporto al tempo. Per dirla meglio, tanto più due specie sono distanti geneticamente tra loro, tanto più l'antenato comune deve essere stato antico; viceversa, se due specie sono prossime sotto il profilo genetico devono aver avuto un antenato comune piuttosto recente. Di più, su queste basi i tempi di divergenza o, per meglio dire, di «coalescenza» possono essere stimati e possono fornire delle date sulla base di un calcolo abbastanza semplice. Questo perché, dicevano i biologi molecolari, le distanze genetiche riguardano per lo più una straripante frazione di DNA che può essere considerata neutra (sic) rispetto alle pressioni della selezione naturale. I paleoantropologi allora capirono in che modo l'evoluzione a livello molecolare è per lo più correlata al tempo, in quanto ogni mutazione neutra (e queste interessano la maggior parte del genoma) può fissarsi nel patrimonio genetico delle specie in base a un criterio meramente probabilistico. Questo fa una bella differenza rispetto all'evoluzione visibile attraverso la morfologia, quella a cui tutti in prima battuta pensiamo e che procede (come si sa) con modi e tempi irregolari: accelerando, rallentando o, addirittura, mostrando morfologie a lungo immutate. Le mutazioni che intervengono invece nella componente neutra del genoma, accumulandosi con regolarità, scandiscono il tempo come un... metronomo. Alla luce delle distanze genetiche e dell'orologio molecolare, una delle ipotesi formulate da Charles Darwin (nel trattato sull' Origine dell'uomo, del 1871) si è rivelata fondata. Cioè che le origini dell'uomo fossero da ricercare in Africa, visto che sono proprio le grandi scimmie antropomorfe africane (gorilla e scimpanzé) a essere più simili a noi di quanto non lo siano le antropomorfe asiatiche (orangutan e, ancor meno, gibboni) o via via tutti gli altri Primati. L'orologio molecolare ha pienamente confermato questa intuizione, affiancando a essa un dato quantitativo impensabile ai tempi di Darwin e aggiungendo una valutazione dei tempi di divergenza (come i 5-6 milioni di anni che separano noi dagli scimpanzé): non è poco. Finalmente così i paleoantropologí accettarono quel benedetto orologio molecolare, rassicurati sul fatto che fosse possibile regolarlo a dovere a partire dal «tasso di mutazione», cioè basandosi su una stima del tempo che, probabilisticamente, ogni mutazione impiega prima di fissarsi nel genoma di una data specie. Il problema è che determinare questo tasso di mutazione non è facile per niente, tant'è che i conti non sempre si combinano a dovere con l'evidenza fossile. Ma di questo riparleremo nelle prossime pagine. | << | < | > | >> |Pagina 196In questa luce, a me piace pensare alla comparsa di Homo sapiens momento davvero cruciale per spiegarci in che modo siamo venuti al mondo come al risultato di una vera e propria «rivoluzione ontogenetica» (come spiego più diffusamente nel mio Il grande racconto dell'evoluzione umana, 2013). In altre parole, penso che con la comparsa della nostra specie sia intervenuto un drastico mutamento, che si sarebbe verificato in seno a una piccola popolazione, isolata dalle altre, nella quale si è realizzata una combinazione genetica che potremmo definire improbabile. Sì, improbabile. Lo dimostra il fatto che tutte le altre linee evolutive del genere Homo quella dei Neanderthal, ad esempio, tanto per fare il caso più vistoso hanno seguito traiettorie di espansione encefalica che non (sottolineo: non) sono state accompagnate da una riconfigurazione dell'architettura cranica, come invece è capitato a noi. Con la speciazione di Homo sapiens, si è dunque verificata una rivoluzione nel processo di accrescimento e sviluppo, con i geni regolatori mutati che controllano ora un nuovo modello morfologico, per il quale si raggiunge una più efficiente armonia tra espansione encefalica e ossificazione del cranio.Osserviamo allora che la nuova specie umana si caratterizza anche per un rapido successo adattativo e demografico, intraprendendo quasi subito una diffusione prima continentale e poi planetaria, che la porterà a contatto con popolazioni arcaiche un po' dappertutto prima in Africa e poi in Eurasia, prima lungo latitudini prossime alla fascia tropicale e poi sempre più a nord e a contribuire, più o meno indirettamente, all'estinzione delle altre specie umane: i Neanderthal in Europa e nel Vicino Oriente, i Denisoviani in Asia continentale, gli ultimi Homo erectus in Indonesia o il piccolo popolo dell'Isola di Flores, ancora più in là. Oltre a diffondersi in Africa e in Eurasia, questa volta l'uomo arriverà fino in Australia e, in seguito, passando dall'area di Bering, si diffonderà anche nelle Americhe. Pur rallentato da altre popolazioni umane e dai fattori geografici, biologici ed ecologici, Homo sapiens si diffonde a un ritmo incalzante. Nel corso della successiva storia naturale della specie, la tecnologia e la cultura hanno costituito il veicolo di una distribuzione geografica pressoché ubiquitaria, attenuando le pressioni ambientali intorno a noi e favorendo l'adattamento di un'unica specie (le altre sono ormai tutte estinte) ai contesti più disparati e abitabili: dalle pianure alle alte montagne, nelle foreste tropicali come nei deserti più aridi, in prossimità dei ghiacciai circumpolari o ingabbiati in affollate metropoli, con tanto di smog e di stress. Tutti gli uomini che siano neri, gialli o bianchi vanno dunque considerati come l'espressione di un unico fenomeno biologico. | << | < | > | >> |Pagina 216Le razze umane non esistonoOltre due secoli fa, gli antropologi si misero al lavoro nel tentativo di comprendere l'inestricabile intreccio della variabilità umana. Presto si affermò una sorta di legittimazione scientifica del razzismo, che ha preso forme di volta in volta condizionate dal clima culturale e socioeconomico dell'epoca. Oggi, fortunatamente, le cose non stanno più così. Il «peccato originale» fu commesso da un grande naturalista francese del '700: Georges-Louis Leclerc de Buffon (1707-1788). Sembra infatti che sia stato proprio lui a importare in biologia il termine race, derivato da un'antica parola in uso all'epoca nel mondo degli allevatori di cavalli: haraz. Con il passare del tempo, la parola razza divenne sempre più usata in ambito zootecnico per indicare le varietà di animali domestici. Da un certo punto di vista, fu l'inizio di una sequela di equivoci, per i quali la scelta di Buffon fu inconsapevolmente cruciale, soprattutto nel momento in cui il termine passò da un contesto di selezione artificiale a quello delle dinamiche evoluzionistiche dettate dai tempi e dai modi della selezione naturale. Quest'ultima svolge un ruolo simile a quello della selezione artificiale operata dagli allevatori, ma non identico. In natura non c'è una mente umana a pilotare gli incroci, l'ambiente non segue alcuno standard né si preoccupa delle nostre classificazioni. In natura, piuttosto sotto l'effetto di pressioni selettive dettate da contesti ecologici diversi popolazioni di una specie con scarse possibilità di movimento e/o di flusso genico tendono, nel tempo, a formare varietà geografiche più o meno isolate tra loro. Non è certo questo il caso della specie umana, tanto meno in tempi recenti. Tuttavia l'uso del termine «razza», ancorché inappropriato, ha pervaso la storia dell'antropologia per un paio di secoli. All'inizio, in pieno Illuminismo, era semplicemente un modo per denotare gruppi di popolazioni accomunate da un certo numero di caratteristiche. Più tardi però, soprattutto nell'800 positivista, l'uso del termine divenne sempre più tipologico. E facilmente la tipologia divenne gerarchia, portando a distinguere fra razze superiori e razze inferiori. Peraltro, la conoscenza del diverso, dell'esotico, proprio nel corso dell'800 raggiungeva livelli mai visti in precedenza, con viaggi di esplorazione in terre lontane e fra popoli altri. Il concetto di razza divenne così un mezzo per catalogare la diversità al servizio delle stesse società che promulgavano le classificazioni, legittimando così il colonialismo, lo schiavismo e varie forme di persecuzione. Alla fine, dal razzismo colonialista dell'800 alle degenerazioni del '900, come il nazismo, i campi di sterminio e altri deliri totalitari, il passo è stato breve. Da questo punto di vista, gli antropologi portano la responsabilità di essersi prestati con le loro classificazioni ad avallare il razzismo. Senza rendersi conto che il programma di ricerca dell'antropologia razziale era fallito in partenza. In effetti, proprio nel tentativo di mettere ordine proliferavano invece le classificazioni più diverse (tanto che ciascun antropologo aveva la sua) e, dunque, spadroneggiava il disordine. Un vero fallimento, e con effetti disastrosi per di più. Va anche detto, però, che lo fecero insieme a buona parte della comunità scientifica e intellettuale del loro tempo. Se consultiamo, per esempio, la lista dei dieci scienziati italiani che nel 1938 firmarono il famigerato Manifesto della razza, solo due erano antropologi (uno dei due, Guido Landra, in vista di una carriera accademica traballante, aveva pensato di «buttarsi in politica»); c'erano anche tre medici, due zoologi, un fisiologo, uno psichiatra e un demografo. Così stanno le cose, così andò.
Oggi, da decenni ormai, la vediamo diversamente e il concetto di razza non
trova più posto in antropologia.
|