|
|
|
|
|
|
|
| << | < | > | >> |Indice
9 Ringraziamenti
11 Condizioni d'ingaggio
Parte prima - L'ARGOMENTO BIOLOGICO
23 1. Vincoli interni: che cosa ci dice la nuova biologia
La selèzione naturale è reale, ovviamente (quando opportunamente definita), 24
Unidimensionalità, 26
Genetica della cesta di semi, 30
Vincoli interni e filtri: "evo-devo", 32
Qualche (indebita) perplessità sulla rivoluzione evo-devo, 36
In sintesi, che cosa abbiamo imparato dall'evo-devo, 37
Altro ancora, ma sempre dello stesso genere, 39
Chaperone, 43
Splicing alternativo, 44
Last but not least: spinta molecolare e conversione genetica, 45
In conclusione di capitolo, 47
49 2. Genomi, reti, moduli e altre cose complesse
Reti di regolazione dei geni, 49
Entrenchment (radicamento generativo), 51
Robustezza, 51
I geni master sono 'padroni", 54
Moduli dello sviluppo, 56
Coordinamento, 60
Esplosioni morfogenetiche, 61
Plasticità e la (non)transitività della fitness, 63
Conclusioni e una nota di cautela, 66
69 3. Molti vincoli, molti ambienti
Selezione senza adattamento, 69
Vincoli sulla selezionabilità e l'ascesa della contingenza, 73
Batteri, zuccheri e vicoli chiusi, 73
Qualche insegnamento da questo caso paradigmatico ultrasemplice, 75
Nuovi tipi di ambienti, 77
Epigenetica e imprinting, 78
Geni che saltano e trasferimenti "orizzontali" di geni, 81
Intercambiabilità fra reazioni a perturbazioni esterne e interne, 84
Conclusioni per questo capitolo, 86
87 4. Il ritorno delle leggi della forma
Un po' di storia, 91
La "quarta dimensione" dei sistemi viventi, 95
Nativismo non-genomico, 97
Altri esempi, 99
La materia grigia e bianca del cervello, 99
Invarianti della locomozione animale, 99
La fisica del canto degli uccelli, 100
Le foglie perfette, 101
Strategie ottimali di procacciamento del cibo: le api, 102
Il colpo d'ala perfetto, 104
La vespa che pilota il suo zombie, 107
Qualche questione relativa all'evoluzione, 108
Un lungo tunnel alla fine della luce, 109
In chiusura di capitolo, 109
Parte seconda - LA SITUAZIONE CONCETTUALE
115 5. Che tipo di teoria è la teoria della selezione naturale?
Introduzione, 115
La selezione naturale come scatola nera, 118
La teoria del condizionamento operante (CO) come scatola nera, 119
Vincoli proprietari: (1) Iteratività, 121
Vincoli proprietari: (2) Ambientalismo, 121
Vincoli proprietari: (3) Gradualismo, 123
Vincoli proprietari: (4) Monotonicità, 125
Vincoli proprietari: (5) Località, 127
Vincoli proprietari: (6) Non-mentalità, 128
Dentro le scatole, 130
Il meccanismo dell'apprendimento secondo la teoria del condizionamento operante, 131
Il meccanismo della selezione secondo TE, 133
135 6. Molti sono i chiamati, ma pochi gli eletti: il problema della "selezione-per"
"Free rider", 136
Avatar del problema della "selezione-per": telelogia, 140
Avatar del problema della "selezione-per", continuazione:
"Che cosa viene appreso?" (Il problema dello "stimolo effettivo"), 142
Avatar del problema della `selezione-per", continuazione:
"Che cosa viene appreso?" (La "risposta effettiva"), 145
Avatar del problema della "selezione-per", continuazione:
la naturalizzazione del contenuto, 148
Un riepilogo provvisorio e prospettive, 151
Da dove arrivano i problemi della "selezione-per"?, 153
Sì, ma che cosa ha a che fare tutto questo con il fatto che la selezione naturale
sia o meno il meccanismo dell'evoluzione dei fenotipi?, 154
160 7. Nessuna via d'uscita? Qualche risposta al problema della "selezione-per"
Ricapitolazione in breve, 161
Prima opzione: dare una possibilità a Madre Natura, 162
Seconda opzione: leggi della selezione, 168
La fallacia della mela svizzera, 169
Terza opzione: il crivello di Sober, 175
Quarta opzione: forse l'adattamentismo non è una teoria, in fin dei conti;
forse è solo uno schema di teoria, 179
Allora che tipo di teoria è la teoria della selezione naturale?, 180
Conclusione retorica, 185
189 8. Il bestiolino ha perso la sua nicchia ecologica? O è stato il contrario?
La corrispondenza raffinata, 191
Ambiente contro nicchia ecologica, 192
202 9. Riepilogo e postludio
215 Appendice
"Quel" genere di darwinisti in psicologia e nella filosofia della mente, 215
Da Boltzmann a W.V.O. Quine, 215
Psicologia evoluzionistica (1988-2009), 218
228 Aggiornamento per l'edizione italiana
Quando piove sul bagnato: recenti convalide della nostra critica al neodarwinismo
(Massimo Piattelli Palmarini)
Freschi di stampa (ottobre 2009 - gennaio 2010), 229
Appena un po' meno freschi, ma non meno significativi, 230
Altri alleati di tutto rispetto, 231
Un dialogo amichevole con Telmo Pievani, 233
Due commentini in chiusura, 235
237 Bibliografia
251 Indice analitico
|
| << | < | > | >> |Pagina 11Condizioni d'ingaggioQuesto non è un libro su Dio; non è un libro sul "disegno intelligente"; non è un libro sul creazionismo. Nessuno di noi ha a che vedere con queste cose. Ci è sembrato opportuno metterlo ben in chiaro subito, perché la nostra affermazione principale, nelle pagine che seguono, sarà che c'è qualcosa di sbagliato - molto probabilmente di fatalmente sbagliato - nella teoria della selezione naturale; e ci rendiamo conto che, persino fra quelli che non sono nemmeno molto sicuri di che cosa sia, l'adesione al darwinismo è diventata una cartina di tornasole per stabilire chi possiede una concezione del mondo "realmente scientifica", e chi no. "Bisogna scegliere fra fede in Dio e fede in Darwin ; e se si vuol essere umanisti laici, meglio optare per la seconda." Così ci dicono. Non siamo affatto convinti che queste due opzioni esauriscano tutte quelle possibili. Ma vogliamo decisamente aderire all'albo degli umanisti laici. In effetti entrambi ci proclamiamo atei - completamente, ufficialmente, fino all'osso e irriducibilmente atei. Perciò cerchiamo spiegazioni esclusivamente naturalistiche dei fatti dell'evoluzione, anche se pensiamo che si dimostreranno molto complesse, come spesso sono le spiegazioni scientifiche. Siamo convinti che l'evoluzione sia un processo totalmente meccanico e siamo convinti che questo escluda non solo cause divine ma anche cause finali, élan vital, entelechie, interventi di alieni extraterrestri e altre cose simili. Il che è, in linea generale, coerente con lo spirito con cui Darwin affrontava il problema dell'evoluzione. Siamo felici - almeno da questo punto di vista - di essere dalla parte di Darwin. Tuttavia, questo libro è soprattutto un lavoro di critica; prevalentemente parla di quello che pensiamo sia sbagliato nel darwinismo. Verso la fine, muoveremo qualche passo nella direzione in cui crediamo possa trovarsi un'alternativa percorribile, ma saranno passi ancora molto vaghi. In realtà, non sappiamo molto bene come funzioni l'evoluzione; non lo sapeva neanche Darwin, e non lo sa esattamente (per quel che possiamo stabilire) nessun altro. "Sono necessarie ulteriori ricerche", come si usa dire. Può darsi che siano necessari secoli di ulteriori ricerche. Forse vi state chiedendo se valga ancora la pena di scrivere una critica del programma darwinista classico. Buoni amici sperimentalisti, che lavorano nel campo della biologia per così dire "umida", ci dicono che nessuno di loro è più "quel tipo" di darwinista; nessuno che lavori nella biologia strutturale è un genuino adattamentista. (Alcune delle ragio- ni per cui non lo sono saranno analizzate nella Parte prima.) Ci fa piacere sapere di questi riallineamenti, ma dubitiamo che siano la norma nella biologia in generale (si pensi, per esempio, alle attuali elaborazioni di modelli matematici per la selezione naturale ottimale). Certo non sono la norma per l'opinione informata in campi in cui ciascuno di noi ha lavorato, come la filosofia della mente, la semantica del linguaggio naturale, la teoria della sintassi, le teorie del giudizio e della decisione, la pragmatica e la psicolinguistica. In tutte queste discipline il neo-darwinismo è assunto come un assioma: non viene mai, letteralmente, messo in questione (si veda l'Appendice). Una concezione che sembri contraddirlo, direttamente o per implicazione, è ipso facto rifiutata, per quanto plausibile possa sembrare. Interi dipartimenti, riviste e centri di ricerca operano secondo questo principio. Di conseguenza il darwinismo sociale cresce rigoglioso, come il darwinismo epistemologico, il darwinismo psicologico, l'etica evoluzionistica e, il cielo ci scampi, l'estetica evoluzionistica. Se volete vedere i loro monumenti, date uno sguardo alle pagine scientifiche dei quotidiani. Abbiamo dedicato entrambi energie e inchiostro per controbattere alcune delle più famose fra queste derive neo-darwiniste, ma pensiamo che sia necessario estirpare l'albero dalle radici; dimostrare che la teoria di Darwin della selezione naturale ha delle falle fatali. Questo è l'obiettivo del libro. Nel corso di queste pagine, cederemo un po' alla tendenza alla digressione. In realtà, la nostra critica del darwinismo solleva questioni collaterali che non possiamo fare a meno di discutere. Perciò ci siamo permessi qualche deviazione che pensiamo sia interessante. Abbiamo comunque una buona scusa: molte questioni che a tutta prima sembrerebbero trasversali rispetto alle nostre finalità, una volta esaminate da vicino, si rivelano invece molto pertinenti. Ogni tanto qualcuno ci chiede se siamo davvero convinti di aver trovato delle "falle fatali" in un corpo teorico che, per così tanto tempo, è stato al centro del consenso scientifico. Ci viene ricordato che la hybris è un peccato e che è bene tenerla a freno. Rispondiamo che, se obiezioni come quelle che solleveremo contro il darvinismo non sono state notate prima, in parte è perché sono finite negli interstizi. Ma è ora di fare un po' d'ordine e riportarle alla luce. Per esempio, seguiremo una linea di argomentazione che procede in questo modo: al cuore delle teorie adattamentiste dell'evoluzione si fa confusione fra (i) l'asserzione che l'evoluzione è un processo in cui vengono selezionati organismi con tratti adattativi e (ii) l'asserzione che l'evoluzione è un processo in cui gli organismi vengono selezionati per i loro tratti adattativi. Sosterremo che il darwinismo è costretto a inferire (ii) da (i); che questa inferenza non è valida (si tratta di quella che i filosofi chiamano "fallacia intensionale"); e che non c'è modo di riparare il danno in modo coerente con il naturalismo, che consideriamo terreno comune. Chiarire tutto questo sarà uno degli obiettivi principali del libro. | << | < | > | >> |Pagina 15Veniamo ora a come abbiamo organizzato la nostra discussione.La Parte prima è dedicata alla biologia, alle ricerche e alle riflessioni più recenti in questo campo. I Capitoli 1, 2 e 3 riassumono un gran numero di fatti nuovi e di nuovi meccanismi non selettivi che sono stati scoperti nella biologia in senso stretto. Spiegano perché i nostri amici biologi non sono più "quel" tipo di darwiniani. Il Capitolo 4 offre un compendio di un altro genere di fatti e di spiegazioni che sono ormai comuni in biologia ma sono estranei alla teoria neo-darwiniana standard dell'evoluzione. In sostanza, riferiamo di casi in cui nei sistemi biologici sono stati trovati strutture e processi ottimali. Sono ottimizzazioni che si presentano in natura, con tutta probabilità determinate dalle leggi della fisica, della chimica e dell'auto-organizzazione spontanea. Pensiamo che in un futuro non molto lontano verranno chiariti altri processi di auto-organizzazione da parte di forze collettive autocatalitiche. Chiaramente, e per ragioni che esamineremo in dettaglio in quel capitolo, non sono l'esito di un filtro esercitato dalla selezione naturale su variazioni generate a caso. La Parte seconda considera le basi logiche e concettuali della teoria della selezione naturale. Il Capitolo 5 espone i molti modi in cui la teoria dell'apprendimento e la teoria evoluzionistica neo-darwinista sono simili, sia nella loro architettura generale, sia per molti dettagli. In particolare, ciascuna è legata a un modello articolato in "generazione e filtraggio" dei fenomeni che si ripropone di spiegare; e ciascuna sostiene che, in prima approssimazione, il generatore in questione è casuale, aleatorio, e il filtro in questione è esogeno. Questi postulati si sono dimostrati insostenibili nella spiegazione dell'apprendimento, e per motivi che sembrano applicarsi parimenti alle spiegazioni basate sulla selezione naturale. L'impostazione "da scienza cognitiva" della psicologia ha in gran parte sostituito la teoria dell'apprendimento, negli ultimi decenni, e ha evidenziato il ruolo dei vincoli endogeni nella formazione dei repertori comportamentali appresi. Con buona pace di Skinner, quel che succede nell'apprendimento non è plausibile si possa rappresentare come il filtraggio esogeno di comportamenti che in prima istanza sono prodotti a caso. Come avremo visto nella Parte prima, esiste un corpus molto persuasivo, e in continua crescita, di prove fattuali che fanno pensare a qualcosa di molto simile nel caso dell'evoluzione dei fenotipi: pensiamo che i darwinisti, come gli skinneriani, abbiamo sovrastimato il ruolo della generazione casuale e del filtraggio esogeno nella formazione dei fenotipi. I Capitoli 6 e 7 si occupano di come nelle spiegazioni adattamentiste dei meccanismi della selezione naturale sorgano quei famigerati problemi di intensionalità cui accennavamo un momento fa. Cominciamo con il fenomeno dei tratti clandestini (in italiano si dice piuttosto "i portoghesi"), cioè tratti fenotipici che viaggiano solidali con, e al seguito di, tratti selezionati (il fenomeno noto come free riding nella teoria dei patti sociali). Sono tratti fenotipici inizialmente neutri sotto il profilo adattativo che vengono trascinati da altri tratti, selezionati, questi, perché influenzano positivamente la fitness. Le discussioni sul free riding si sono diffuse nella letteratura biologica dopo un famoso saggio di Gould e Lewontin (1979) (in realtà, il fenomeno è stato riconosciuto dallo stesso Darwin) e si ammette generalmente che costituisca un'eccezione per le spiegazioni strettamente adattamentiste dell'evoluzione. Ma quasi tutti pensano che si tratti di un'eccezione relativamente marginale: un'eccezione che si possa ammettere senza dover coerentemente rinunciare a pensare che l'evoluzione dei fenotipi sia influenzata in primo luogo dalla selezione esogena. Dimostreremo, però, che così proprio non si colgono le implicazioni del free riding e dei fenomeni correlati per le teorie dell'evoluzione deì fenotipi. I darwinisti sono tormentati dal free riding perché non si sono resi conto dell'intensionalità della nozione di "selezione-per" e di altre simili; e quando si richiama la loro attenzione sul problema, non sanno minimamente che pesci pigliare. Pensiamo che questa situazione abbia generato quell'esercito di fantasmi da cui sono sempre più infestate le spiegazioni darwiniste dell'evoluzione: Madre Natura, geni egoisti, memi imperialisti e altro ancora, esempi assai comuni nella letteratura corrente. Ma le radici dei problemi dei free rider risalgono a Darwin, e alla sua passione per la supposta analogia fra il modo in cui la selezione naturale manipola i fenotipi e il modo in cui li manipolano gli allevatori. Sotto questo aspetto Darwin non è stato abbastanza colpito dal fatto che gli allevatori hanno una mente: agiscono seguendo le loro convinzioni, i loro desideri, le loro intenzioni e così via; mentre, ovviamente, nessuna di queste cose vale nel caso della selezione naturale. Sarebbe davvero stupefacente, alla luce di questa differenza, se i processi della selezione artificiale fossero modelli affidabili per quelli della selezione naturale. Il Capitolo 8 risponde a quello che è forse l'argomento più forte a sostegno della selezione naturale come meccanismo principale dell'evoluzione dei fenotipi, cioè che nessuna alternativa può fornire un resoconto naturalistico del "raffinato adattamento" dei viventi alle loro ecologie. Pensiamo che questo argomento, anche se onnipresente nella letteratura, sia fallace. E questo capitolo spiega perché lo pensiamo. Nel Capitolo 8 ci soffermiamo su come i problemi relativi alla selezione naturale interagiscano con questioni più generali relative alla spiegazione scientifica. Alcune spiegazioni empiriche caratterizzano i loro domini simultaneamente a più livelli ontologici. Le spiegazioni storiche, per esempio, sono splendidamente "a molti livelli". Una spiegazione del perché Napoleone fece quello che fece a Waterloo può fare appello simultaneamente alla sua età, alla sua formazione, alla sua classe sociale e al suo tipo di personalità, per non parlare della sua esperienza militare passata, delle sue condizioni psicologiche, del tempo atmosferico e di quanta caffeina c'era nel caffè che prese quel mattino. Ci sono invece teorie "a un solo livello", delle quali la meccanica newtoniana è forse l'esempio estremo. Gli individui contemplati nel dominio di quella teoria si immagina siano costituiti esclusivamente da masse puntiformi e tutto quello che la teoria "sa di essi" (i parametri in base ai quali si applicano le leggi della teoria) sono le loro posizioni, le loro velocità e le forze che su di essi agiscono. Così la meccanica newtoniana fa astrazione dal colore delle cose, dalle loro storie individuali, da chi eventualmente le possieda e così via. Spesso si osserva che gli individui considerati dalle scienze di base sono più simili fra loro di quelli che prendono in esame le scienze speciali. Mentre le particelle newtoniane si differenziano solo per la loro massa, la loro posizione o la loro velocità, organismi, fenotipi ed ecologie possono essere (e sono) diversi in moltissimi modi. Si veda Ellis (2002) per una discussione interessante in proposito. Molti suppongono (ma, secondo noi, erroneamente) che le teorie dell'evoluzione siano fondamentalmente del tipo a un solo livello. Le uniche relazioni che ammettono sono quelle che valgono fra oggetti (e/o eventi) al macrolivello: organismi da una parte e le loro ecologie dall'altra. Le spiegazioni offerte dalle teorie di questo genere specificano come le interazioni causali fra gli organismi e le ecologie producano cambiamenti della fitness nei primi. Da questo punto di vista, Darwin e Newton seguono le stesse linee di lavoro. Il Capitolo 9, quello conclusivo, considera quanto sia plausibile che l'evoluzione possa essere spiegata da una teoria a un solo livello. Fra le morali principali che questo libro vuole trarre, vi è che i processi di alterazione fenotipica non hanno molto di interessante in comune in quanto tali. Se è vero, ne seguono alcune cose importanti: dato che non esiste un unico meccanismo di fissazione dei fenotipi, c'è un senso importante in cui non esiste un "livello" di spiegazione evoluzionistica, e non ci possono essere teorie generali dell'evoluzione. La storia dell'evoluzione dei fenotipi, invece, non appartiene alla biologia ma alla storia naturale; e la storia, naturale o meno, è il luogo per eccellenza delle spiegazioni che non sono conformi al paradigma newtoniano. Abbiamo incluso anche un'appendice di citazioni che mostrano quanto le versioni "dure" dell'adattamentismo si siano infiltrate in campi adiacenti alla biologia, fra cui la filosofia, la psicologia e la semantica. Questa dunque la struttura generale. Chiudiamo queste note di prefazione con una breve omelia. Più di uno dei nostri colleghi ci ha detto che, anche se Darwin aveva sostanzialmente torto a sostenere che la selezione naturale è il meccanismo dell'evoluzione, comunque non dovremmo dirlo. Non, comunque, in pubblico. Per quanto involontariamente, comportarsi in questo modo significherebbe schierarsi con le Forze dell'Oscurità, il cui obiettivo è portare discredito alla Scienza. Beh, non siamo d'accordo. Pensiamo che il modo per mettere in difficoltà le Forze dell'Oscurità sia seguire le argomentazioni dovunque possono condurre, e così facendo diffondere il massimo possibile di luce. Quel che rende oscure le Forze dell'Oscurità è che non sono disposte a fare una cosa del genere. Quel che rende scientifica la Scienza è, invece, che è dispostissima. | << | < | > | >> |Pagina 24La selezione naturale è reale, ovviamente (quando opportunamente definita)Non c'è dubbio che mutamenti degli equilibri (cioè variazioni delle frequenze relative dei tipi fenotipici all'interno di una stessa popolazione e fra popolazioni diverse) avvengono continuamente, sulla terraferma, nei mari, nei laghi, nei fiumi di tutto il pianeta. Si verificano anche nei nostri organismi: avvengono alterazioni delle cellule epiteliali (della pelle), delle cellule pancreatiche, dei linfociti (globuli bianchi del sangue), di neuroni e sinapsi, anche nel nostro corpo mentre scriviamo queste righe, nel vostro mentre le leggete. Questi mutamenti non hanno sosta e si verificano sulla Terra da centinaia di milioni di anni. Poi reti di relazioni di predazione, commensalismo (condivisione del cibo), competizione e migrazione si intrecciano con questi mutamenti e modificano, sul lungo periodo, la nostra struttura e quella dei nostri ecosistemi. Le distribuzioni di tratti biologici e comportamentali nelle popolazioni che vediamo oggi sono il risultato di questi processi, anche se non solo e non principalmente di questi (ipotizzando che si possa definire una misura affidabile - una metrica sensata - per valutazioni probabilistiche di questo genere, un argomento su cui torneremo). È convinzione comune che le distribuzioni di tratti fenotipici nelle popolazioni varino lentamente ma costantemente nel tempo. Detto questo, però, bisogna sottolineare che questi equilibri mobili non spiegano la distribuzione dei fenotipi; sono invece fra i fenomeni che le teorie dell'evoluzione dovrebbero spiegare. Oggi i biologi hanno buoni motivi per credere che la selezione fra varianti di poco conto, e generate a caso, di tratti fenotipici non possa proprio spiegare la comparsa di nuove forme di vita. Non aiuta ipotizzare che l'evoluzione abbia luogo nell'arco di periodi di tempo molto, molto lunghi, se (come pensiamo) fattori endogeni e regolazioni genetiche su molti livelli hanno un ruolo essenziale nel determinare le opzioni fenotipiche fra cui le variabili ambientali possono scegliere. Contrariamente all'opinione tradizionale, bisogna sottolineare che la selezione naturale fra tratti generati in modo casuale non può essere il principio base dell'evoluzione. Devono esserci invece vincoli endogeni forti, spesso decisivi, e schiere di regolazioni delle opzioni fenotipiche su cui opera la selezione esogena. In questo senso, per noi la selezione naturale è come accordare il pianoforte, non come comporre delle melodie. Questa è la nostra storia, e pensiamo che sia la storia che la biologia moderna racconta quando viene presentata nel modo giusto. A questa storia ci atterremo in tutto quello che segue. Pensiamo (e difenderemo questa posizione nei capitoli seguenti) che esistano argomenti a priori convincenti che lo dimostrano. Per il momento, però, ammettiamo che spesso è molto difficile prevedere gli effetti dell'applicazione di un processo di selezione a una popolazione di tratti generati casualmente. Anche minime variazioni nelle frequenze iniziali, nei tassi di mutazione casuale e nei coefficienti di selezione possono portare a nuovi equilibri drasticamente differenti. Questo capitolo presenta in sintesi una panoramica dei meccanismi specifici la cui scoperta rende (almeno in certi casi) palesemente sbagliata la teoria gradualista/adattamentista della selezione naturale, perché i nuovi tratti fenotipici non sono generati a caso (come avverrebbe se le mutazioni che esprimono fossero indipendenti) o perché l'adattamento all'ecologia ha un ruolo solo secondario nella determinazione dei fenotipi, o per entrambe queste ragioni. | << | < | > | >> |Pagina 26UnidimensionalitàLa dottrina tradizionale della selezione naturale (SN) è a una sola dimensione. Si assume che la struttura ecologica sia la spiegazione par excellence della struttura fenotipica; il contributo delle fonti interne (endogene) di variazione e dei vincoli interni è considerato, nel migliore dei casi, marginale. Questo spinge a pensare che, ai fini di una spiegazione evoluzionistica, si possa fare astrazione dal carattere delle connessioni fra i geni e le loro espressioni fenotipiche, nonché dal carattere del genoma stesso. Tutta questa struttura interna è vista come ampiamente irrilevante per spiegare il corso dell'evoluzione: la selezione naturale troverà le sue soluzioni indipendentemente dai particolari genetici. L'esempio più chiaro e più autorevole di questa posizione si trova in Ernst Mayr , uno dei principali artefici della sintesi moderna (cioè della fusione del darwinismo classico con la genetica, a partire dai primi decenni del XX secolo). Citiamo quella che sembra, col senno di poi, e sullo sfondo della rivoluzione evo-devo (vedi oltre, in questo capitolo) un'affermazione davvero stupefacente:
Gran parte di quello che abbiamo imparato sulla fisiologia del
gene rende evidente che la ricerca di geni omologhi è del tutto futile, tranne
che per parenti molto stretti. Se esiste solo
una soluzione efficiente per una determinata esigenza funzionale, complessi di
geni molto diversi fra loro troveranno la
stessa soluzione, per quanto diverso sia il percorso grazie al
quale la raggiungono. Il detto "tutte le strade portano a Roma" è vero
nell'evoluzione così come nelle faccende di tutti i giorni.
Mayr, 1963, p. 609
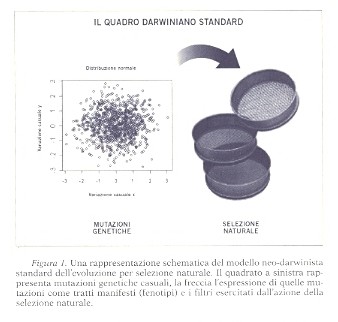
Come vedremo fra breve, sia i casi frequenti di conservazione degli stessi geni master nell'arco di centinaia di milioni di anni, sia tutto il campo dell'evo-devo raccontano una storia del tutto diversa. La rivoluzione evo-devo ci dice che nulla nell'evoluzione ha senso se non alla luce della biologia dello sviluppo - l'affermazione di Mayr è sintomatica del potere illimitato attribuito alla selezione naturale. Un caso paradigmatico è quello della formazione dell'occhio in specie distanti, che si suppone abbia avuto luogo in modo convergente e indipendente molte volte nel corso dell'evoluzione (almeno cinque volte, e forse molte di più). Ma allora, con la scoperta degli stessi geni master per lo sviluppo dell'occhio (in particolare Pax3, Pax2, Pax6 e Dach) in classi e specie molto distanti (dal riccio di mare, dove i geni restano inespressi, alle meduse, ai moscerini della frutta, ai vertebrati) la scena evolutiva è cambiata moltissimo (per dati stupefacenti e considerazioni relative si veda Sherman, 2007). Come vedremo nei capitoli successivi, il quadro che ricaviamo dalla sintesi moderna è simile a quello che la teoria comportamentista dell'apprendimento propone in psicologia: un generatore casuale di diversità, che si proietta su fenotipi che, a loro volta, soddisfano a quanto richiesto dai filtri ambientali. Lo schema può essere visualizzato come nella Figura 1.
Senza alcun dubbio le considerazioni relative alla freccia nel diagramma sono sempre state ritenute importanti nel mondo dei sistemi biologici reali, ma sono state reputate ampiamente irrilevanti per la teoria dell'evoluzione. Come testimonia la citazione da Mayr, si postulava che la teoria della selezione naturale potesse fare astrazione dai dettagli dell'organizzazione genetica e da quelli dei processi di sviluppo; questi ultimi lasciati alla cura degli embriologi. Fondamentalmente, nella nuova sintesi, l'evoluzione convergente era considerata onnipresente, anziché occasionale, e le variabili endogene erano trattate come casuali; si postulava che tutto il lavoro fosse fatto dalle variabili esogene. Questo è ciò che dà alla teoria il suo carattere unidimensionale. Una sola "grande" freccia. | << | < | > | >> |Pagina 45Last but not least: spinta molecolare e conversione geneticaDeplorando il monopolio tradizionalmente attribuito alla selezione naturale e alla deriva genetica casuale come uniche cause di formazione di nuove specie, Gabriel Dover ha proposto un nuovo meccanismo, denominato molecular drive, spinta molecolare (Dover 1982a, b). In sostanza, la proposta di Dover consisteva in un processo molecolare di turnover (ricambio) interno al genoma, indipendente dalla selezione naturale. Uno schema concertato di fissazione che permette la formazione di forme biologiche nuove "in un modo non previsto dalla genetica classica della selezione naturale e della deriva genetica". Nello spiegare, con gli strumenti che aveva a disposizione agli inizi degli anni ottanta, i possibili dettagli di questi processi molecolari completamente meccanicistici e totalmente interni all'apparato genetico, Dover usava tre parole che sono diventate molto importanti, più di 25 anni dopo: "direzionale", "biased" (polarizzato) e "conversione". In sostanza, Dover sottolinea che tutti i genomi di tutte le specie esaminate, dai moscerini ai vermi agli esseri umani, sono resi più complessi dagli onnipresenti meccanismi genomici del turnover (trasposizione replicativa, inversione, duplicazione) che alimentano quella che chiama "spinta molecolare". L'instabilità ricorrente dei genomi porta a riorganizzazioni e a nuovi stati stabili temporanei. Dover evidenzia che le conseguenze sempre più diffuse della spinta molecolare funzionano esattamente nello stesso modo (errore di campionamento) in cui agisce la deriva genetica stocastica a livello del fenotipo. Oltre alla conversione dei geni (biased e unbiased), a cui torneremo fra breve, esistono trasposizioni, scivolamenti, crossingover (incroci) disuguali di cromosomi e altri processi che garantiscono, nella visione di Dover, che quella che inizialmente è una singola mutazione in un singolo gene in un singolo cromosoma in un singolo individuo può irradiarsi, con il passare delle generazioni, entro tutta una popolazione che si riproduce sessualmente. Questo processo di radiazione alimentato internamente, nel corso del tempo, può aprire per una popolazione percorsi di sviluppo, di riproduzione e di comportamento che in precedenza non le erano accessibili. Nello schema di Dover (si veda il suo libro del 2001, Dear Mr. Darwin) si può pensare che lo stabilirsi di nuove funzioni, amichevoli per l'ambiente, sia dipendente da un'interazione fra molti processi: spinta molecolare, deriva casuale e (sì, anche quella) selezione naturale. Nel suo schema, riassumendo drasticamente, sono attive forze che fondamentalmente sono dovute all'instabilità dei genomi (onnipresenti meccanismi non-mendeliani di turnover), che consentono una comprensione radicalmente più ampia della natura evoluta delle funzioni biologiche. Oggi si sottolinea un aspetto capitale delle prime intuizioni di Dover (e dei suoi dati e dei suoi calcoli): il processo denominato biased gene conversion (BGC). Oggi si osserva, cioè, che questo meccanismo, che si riferisce alla ricombinazione dei geni, con conseguente segregazione e distorsione, guida la fissazione di nuove varianti genetiche (nuovi alleli) indipendentemente da qualsiasi processo selettivo. | << | < | > | >> |Pagina 51RobustezzaUn tratto si dice robusto rispetto a una variabile genetica o ambientale se la variazione dell'uno è solo debolmente correlata a variazioni dell'altra. In altre parole, la robustezza è la persistenza di un tratto di un organismo a fronte di perturbazioni, che siano rumore casuale nello sviluppo, cambiamenti ambientali o genetici. [...] Recensendo su "Science" (2005) un libro sulla robustezza e la possibilità di evolvere scritto da Andreas Wagner, Gregory C. Gibson (William Neal Reynolds Distinguished Professor of Genetics all'Università statale della North Carolina) diceva: La robustezza deve comportare interazioni genetiche non additive, ma i genetisti quantitativi, per la maggior parte di un secolo, hanno dato per scontato che sia solo la componente additiva della variazione genetica che risponde alla selezione. Di conseguenza, ci troviamo di fronte all'osservazione che i sistemi biologici sono pervasivamente robusti ma troviamo difficile spiegare esattamente come evolvano per diventare tali. Gibson, 2005, p. 237, corsivo nostro Musica per le nostre orecchie, in questa recensione Gibson aggiunge che "[questo libro] dà un contributo significativo alla concezione emergente per cui la selezione sarebbe solo una, e forse nemmeno la più fondamentale, fra le fonti di ordine biologico". Lasciateci sottolineare "concezione emergente" e "nemmeno la più fondamentale". Dicevamo, appunto, musica per le nostre orecchie. Lo stesso Darwin aveva esplicitamente ammesso che la selezione naturale non è l'unico meccanismo dell'evoluzione, ma val la pena sottolineare che oggi, come dice Gibson (sia pur prudentemente aggiungendo un "forse"), "non è nemmeno il più fondamentale". Vogliamo proseguire su questa strada e concludere che questi livelli molteplici di vincoli interni sui fenotipi possibili rendono radicalmente insostenibile l'idea dell'evoluzione come prodotto di una selezione esterna che agisce su variazioni fenotipiche generate in modo casuale. Darwin sosteneva che (per usare un'espressione di Dennett ) i fenotipi "portano informazioni su" le ecologie in cui sono evoluti. Il colore scuro della farfalla ci dice che è evoluta in un'atmosfera piena di smog. Ma ora sembra innegabile che i fenotipi evoluti portino anche informazioni sull'organizzazione interna degli organismi che li possiedono (per esempio, in merito alle loro strutture genotipiche e ontogenetiche). È una questione aperta, empirica e di grande sostanza quanto strettamente questi effetti endogeni vincolino le variazioni fenotipiche su cui opera la selezione esterna. Ci vorrà un po' per scoprirlo, ma, fino a che la domanda non avrà una risposta, non è consigliabile dare per scontata una spiegazione neo-darwinista dell'evoluzione. | << | < | > | >> |Pagina 107La vespa che pilota il suo zombieInfine, un caso (anche qui, fra i molti) in cui la programmazione genetica di un comportamento complesso non lascia alcun dubbio. Si può provare che questi comportamenti sono completamente automatici lungo la sequenza completa, e non appresi. Per farla breve, una particolare specie di vespa (Ampulex compressa) usa un cocktail di veleni per manipolare il comportamento della sua preda, uno scarafaggio. Come in alcune altre specie di vespe solitarie, la vespa femmina paralizza lo scarafaggio senza ucciderlo, e poi lo trasporta nel suo nido e deposita le sue uova nel ventre della preda, in modo che i neonati possano nutrirsi del corpo vivente dello scarafaggio. Quel che è peculiare di questa specie di vespe è il fatto che, mediante due punture consecutive, separate da un intervallo temporale molto preciso, in due parti diverse e scelte con precisione del sistema nervoso dello scarafaggio, la vespa riesce letteralmente a "guidare" nel suo nido già predisposto la preda trasformata in uno zombie. La vespa non deve trascinare fisicamente lo scarafaggio nel suo rifugio, perché può manipolare le antenne della preda, o letteralmente cavalcarla, dirigendola come se fosse un cane al guinzaglio o un cavallo alla briglia (Libersat, 2003). La prima puntura nel torace provoca una paralisi momentanea delle zampe anteriori, che dura qualche minuto, bloccando alcuni comportamenti ma non altri. La seconda puntura, parecchi minuti più tardi, è direttamente sul capo.
Il risultato è che la vespa può afferrare una delle antenne
dello scarafaggio e farlo andare fino al luogo adatto all'ovo-deposizione. Lo
scarafaggio segue la vespa docilmente come
un cane al guinzaglio (Williams, 1942; Fouad et al., 1994).
Pochi giorni più tardi, lo scarafaggio, immobilizzato, funge
da fonte di cibo fresco per la prole della vespa.
Qualche questione relativa all'evoluzione
Questa macabra storia entomologica fa sorgere qualche
questione chiave per l'evoluzione. In un simile comportamento complesso,
sequenziale, rigidamente pre-programmato,
molte cose avrebbero potuto andare storte, in molti modi, in
ciascuno dei suoi passi. La natura biochimica del cocktail di
veleni avrebbe potuto essere molto diversa, risultando o del
tutto inefficace o, per eccesso, letale per la preda. La scelta
del momento e dei punti in cui pungere avrebbe potuto essere sbagliata in molti
modi, per esempio consentendo allo scarafaggio di riprendersi e di uccidere la
vespa, di lui molto più piccola. La vespa avrebbe potuto non "capire" che la
preda può essere guidata al guinzaglio, dopo le due magistrali punture, e
avrebbe potuto tentare di trascinare faticosamente il
corpo piuttosto voluminoso nel suo nido. E via di questo passo. I modi in cui
questa sequenza comportamentale avrebbe
potuto uscire di strada sono in effetti innumerevoli. Neanche il più convinto
fra gli adattamentisti neo-darwinisti suppone che gli antenati della vespa
abbiano tentato alla cieca
tutti i tipi di alternative e che siano state progressivamente
selezionate soluzioni sempre più valide, fino a che non è stata trovata la
soluzione ottimale, che è stata conservata e codificata nei geni. Vero: le vespe
sono in circolazione da moltissimo tempo (circa 400 milioni di anni, forse più),
ma anche questo non è un tempo abbastanza lungo per tentare a
casaccio un numero enorme di soluzioni comportamentali
alternative, con possibilità alternative concepibili a ogni
passo della sequenza comportamentale. E allora? Nessuno
lo sa, al momento. Simili casi di programmi comportamentali innati complessi
(raffinate ragnatele, procacciamento
del cibo nelle api come abbiamo visto prima, e molti altri)
non possono essere spiegati direttamente mediante fattori
ottimizzanti fisico-chimici o geometrici. Ma non possono
essere spiegati nemmeno dall'adattamento gradualistico. È
corretto ammettere che, anche se siamo disposti a scommettere che un giorno si
troverà una spiegazione naturalistica,
per il momento non ne abbiamo nessuna. E se insistiamo
che la selezione naturale è l'unica via da esplorare, non ne
avremo mai una.
Un lungo tunnel alla fine della luce
I 24.000 geni circa nel genoma umano possono arrivare
fino a un certo punto, per assemblare un essere umano. A
dire il vero, come abbiamo visto, ci sono molte regolazioni
genetiche e reti di interazioni, e attrattori morfogenetici, e
modifiche epigenetiche, e
splicing
di geni alternativi e interazioni complesse con molti ambienti. Questi ultimi,
nel caso degli esseri umani, comprendono anche la "cultura". Il
compito di questi relativamente pochi geni sembra spaventoso, anche considerando
il notevole "moltiplicatore" rappresentato dallo
splicing
alternativo (vedi sopra). Fra le molte altre strutture complesse, si devono
produrre decine di milioni di tipi di anticorpi, si devono sviluppare e fissare
10^11 neuroni e 10^13 sinapsi, e circa 90.000 chilometri di vene, arterie e
capillari che vanno collocati in posizioni esatte
in ciascuno dei nostri organismi. Sicuramente sono attivi
molti processi di auto-organizzazione spontanea, a molti livelli. Qui la fisica
dei fenomeni collettivi lascia una firma, in
modi che ancora ci sfuggono. Sia i processi genetici-epigenetici, sia questi
altri processi hanno cause interne. L'idea di
Cherniak di "nativismo non-genomico" (Cherniak et al.,
1999; Cherniak et al., 2004; Cherniak, 2009) è appropriata e,
sembra, ineludibile.
In chiusura di capitolo
I neo-darwinisti sono propensi a dire che la selezione naturale non ottimizza mai, ma si limita a trovare soluzioni localmente soddisfacenti. Dal "bricolage" evoluzionistico di François Jacob al "satisficing" di Maynard Smith e Dennett si mette sempre l'accento su questa considerazione. Per la nostra critica del neo-darwinismo è importante che il problema del trovare soluzioni ottimali a problemi dell'evoluzione mediante setacciamento di candidati generati casualmente sarebbe spesso intrattabile. Ma, come abbiamo appena visto, esistono alcuni casi di soluzioni ottimali (o quasi-ottimali) per problemi di biologia; così, se la selezione naturale non può ottimizzare, deve essere coinvolto qualcosa d'altro. Molto plausibilmente, il "qualcosa d'altro" comprende: fisica, chimica, processi autocatalitici, strutture dissipative e principi di auto-organizzazione, e sicuramente altri fattori che il progresso della scienza al momento opportuno ci rivelerà. La morale qui è una sorta di dilemma per i neo-darwinisti: anche se supponiamo, per amor di discussione, che la selezione naturale operi nel modo in cui il neo-darwinismo canonico sostiene che operi, i suoi gradi di libertà debbono essere fortemente limitati. La piccolissima parte effettivamente occupata dei morfospazi della vita (McKinney e McGhee, 2003; McGhee, 2007; Raup, 1966) è qualcosa che la teoria della selezione naturale non può spiegare. Siamo comprensibilmente meravigliati davanti alla varietà e diversità delle forme della vita, ma è importante sottolineare che, a un livello astratto, quando si riportano in grafico le possibili variazioni continue dei parametri della forma, le forme di vita attuali ed estinte occupano un piccolissimo sottoinsieme di quello che in astratto è possibile. Come abbiamo appena detto, il dilemma resterebbe anche se la teoria della selezione naturale fosse per altro fondamentalmente corretta. Ma il problema è puramente accademico, poiché i capitoli che seguono (nella Parte seconda) dimostreranno che corretta non è. | << | < | > | >> |Pagina 1155. Che tipo di teoria è la teoria della selezione naturale?Introduzione La teoria (neo-)darwiniana dell'evoluzione (per brevità TE) è formata da due parti distinte ma in relazione fra loro: una spiegazione storica della genealogia delle specie (GS) e la teoria della selezione naturale (SN). La tesi di fondo di questo libro è che SN sia irreparabilmente difettosa. Non abbiamo invece alcuna critica da muovere alla genealogia delle specie; è perfettamente possibile - in effetti, del tutto probabile - che GS sia vera anche se SN non lo è. Perciò siamo pronti a concedere, almeno ai fini della discussione che segue, che la maggior parte o tutte le specie siano in relazione fra loro in grazia della loro discendenza storica, magari da un comune progenitore primitivo; e che si possa assumere come regola empirica che, più simili sono i fenotipi di due specie, meno lontano nel tempo sia il primo antenato che hanno in comune. Anche se assumiamo che GS e SN siano indipendenti, non immaginiamo che siano prive di collegamenti. Pensate la genealogia delle specie come un albero (o forse un cespuglio) costituito da nodi e archi (rami); ogni nodo rappresenta una specie e ogni specie è un antenato di tutti i nodi (di livello successivo) che le sono collegati. A questo punto sorge la domanda: com'è successo che la tassonomia delle specie diventasse quello che è? Che cosa determina quali nodi ci sono e quali cammini esistono fra di essi? In particolare, per quali processi una specie progenitrice si differenzia nei suoi discendenti? Queste sono le domande a cui l'adattamentismo di Darwin pretende di dare una risposta. E la risposta che propone è che se, nell'albero genealogico, esiste un cammino a ritroso che porta da A a B, allora A è derivato dalla specie B attraverso un processo di selezione naturale e il cammino da un nodo all'altro corrisponde al dispiegarsi di quel processo. Sosteniamo che questa risposta proprio non è giusta; qualsiasi cosa sia SN, non può essere il meccanismo che genera la tassonomia storica delle specie. Jared Diamond, nella sua Introduzione a Mayr (2001, p. x) osserva che Darwin non si è limitato a presentare "una teoria dell'evoluzione ben elaborata. Cosa di gran lunga più importante, ha proposto anche una teoria della causazione, la teoria della selezione naturale". Beh, se abbiamo ragione, questo è proprio quello che Darwin non ha fatto; o, se preferite, Darwin ha effettivamente proposto un meccanismo causale per il processo di speciazione, ma non era quello giusto. C'è una certa ironia storica in questo, perché la genealogia darwiniana e non la teoria della selezione naturale è stata fonte di tante controversie politiche e teologiche negli ultimi cento e più anni. Per dirla brutalmente: chi non ama il darwinismo ha contestato in modo particolare l'implicazione che nel suo albero di famiglia ci sia un babbuino; più precisamente, non è disposto a riconoscere di avere un antenato (recente) in comune con il babbuino; così non si pone neanche il problema di come la scimmia ancestrale abbia potuto evolvere da una parte negli esseri umani e dall'altra nei babbuini. Questo libro è anti-darwinista, ma (lo ripetiamo) non di quel tipo di anti-darwinismo. Tranquillamente inghiottiamo in famiglia sia il babbuino sia la scimmia progenitrice (e, di questi tempi, ben altre e ben inferiori specie animali), ma non la tesi che la selezione naturale sia il meccanismo della speciazione. L'argomentazione, da cui si conclude che c'è qualcosa di sbagliato nella selezione naturale, è in realtà molto diretta; in una certa misura, è addirittura familiare. Ma non per le discussioni intorno al darwinismo in sé, bensì per i problemi che sorgono in campi adiacenti come la metafisica del riferimento, lo status della teleologia biologica e, soprattutto, la psicologia dell'apprendimento. Mettere a nudo la somiglianza - addirittura l'identità - astratta di queste questioni prima facie eterogenee è uno degli obiettivi principali di quel che segue. Per questo però sarà necessaria un'esposizione un po' idiosincratica di SN. In primo luogo, proponiamo di introdurre SN distinguendo fra (i) la teoria considerata semplicemente come una "scatola nera" (cioè semplicemente una funzione che mette in corrispondenza certi tipi di input con certi tipi di output) e (ii) la spiegazione che la teoria dà dei meccanismi che calcolano quella funzione e dei vincoli sotto i quali vengono eseguiti i calcoli. È un modo un po' eccentrico, diciamo così, di tagliare la torta; ma darà i suoi frutti quando cercheremo di chiarire quali sono secondo noi i problemi di SN. In secondo luogo, vogliamo condurre la nostra esposizione della spiegazione darwiniana dell'evoluzione in parallelo con un'esposizione della teoria di B.F. Skinner dell'apprendimento per condizionamento operante (CO). Alcune somiglianze fra le due teorie sono state ampiamente notate, non ultimo dallo stesso Skinner; ma pensiamo comunque che la portata dell'analogia fra SN e CO sia seriamente sottovalutata e che le implicazioni in genere siano state fraintese. In effetti, le due teorie sono praticamente identiche: propongono sostanzialmente gli stessi meccanismi per calcolare funzioni sostanzialmente simili sotto vincoli sostanzialmente identici. Questo solleva una domanda su cui le precedenti discussioni di SN sono state, ci sembra, decisamente reticenti: si è generalmente concordi, oggi, che la spiegazione skinneriana dell'apprendimento sia morta senza speranza di resurrezione. E allora, se è vero che la teoria di Skinner e quella di Darwin sono variazioni sullo stesso tema, perché le obiezioni che vengono comunemente sollevate alla prima non vengono avanzate anche alla seconda? Se nessuno crede più a Skinner, perché invece tutti credono ancora a Darwin? Sosterremo che la posizione, che mantiene la seconda ma non la prima, non è difendibile. | << | < | > | >> |Pagina 152A questo punto possiamo annunciare la nostra strategia polemica generale: abbiamo aperto il capitolo ricordando l'intuizione di Gould e Lewontin , che una teoria della selezione naturale deve in qualche modo consentire la possibilità di tratti fenotipici che non sono adattamenti. Pensiamo che Gould e Lewontin avessero completamente ragione in proposito, ma pensiamo che sia sfuggito loro un punto più profondo: una volta che il carattere dei problemi di "selezione-per" sia compreso correttamente, diventa evidente che la domanda sollevata dai free rider fenotipici non può avere una risposta nella cornice delle teorie adattamentiste dell'evoluzione. Se è vero, allora l'adattamentismo semplicemente non può fare quello che si presume debba fare una teoria dell'evoluzione: spiegare come i tratti fenotipici siano distribuiti nelle popolazioni di organismi. Detto in modo diverso ma equivalente: la teoria della selezione naturale non può prevedere/spiegare per quali tratti sono selezionati gli individui in una popolazione.L'argomentazione si dipana in tre fasi. Prima suggeriamo una diagnosi: vogliamo chiarire che cosa fa sì che una teoria generi problemi della "selezione-per". Se la diagnosi è giusta, ci porta direttamente a un modo per distinguere i tipi di problemi della "selezione-per" che hanno soluzione da quelli che invece non l'hanno. Infine, sosterremo che (sulla base di assunzioni empiriche plausibili) i problemi della "selezione-per" che affliggono l'adattamentismo sono del tipo non risolubile. Il resto del capitolo coprirà le prime due fasi dell'argomentazione; la terza occuperà il Capitolo 7. | << | < | > | >> |Pagina 180Allora che tipo di teoria è la teoria della selezione naturale?Voce dell'esasperazione: "Ma voi volete proprio dire che le spiegazioni adattamentiste non valgono mai niente; che le storie di selezione non spiegano mai i tratti fenotipici, che siano psicologici o d'altro genere? Vi rendete sicuramente conto che i manuali sono semplicemente pieni di buoni esempi in contrario. Queste spiegazioni da manuale intendono dare, e spesso chiaramente danno, le ragioni per cui i fenotipi sono così come sono; perché ci sono molte popolazioni di t1, ma poche o nessuna popolazione di t2. E allora, che cosa ce ne facciamo dei paradigmi da manuale di spiegazioni adattamentiste se, come dite voi, l'adattamentismo non è vero ma vuoto?". Pensiamo che in effetti ci siano davvero alcune genuine spiegazioni adattamentiste e che quello che sono sia precisamente quello che sembrano essere superficialmente: sono spiegazioni storiche. Molto alla buona, le spiegazioni storiche offrono (non leggi, ma) narrazioni plausibili; narrazioni che intendono articolare la catena causale degli eventi che porta all'evento da spiegare. Le spiegazioni nomologiche riguardano relazioni (metafisicamente necessarie) fra proprietà; le narrazioni storiche riguardano relazioni (causali) fra eventi. Per questo le prime sostengono dei controfattuali, ma le ultime no. Le narrazioni storiche, per quanto ne sappiamo, vanno benissimo; certo spesso sono del tutto persuasive. Ma non sussumono eventi sotto leggi e perciò non sostengono controfattuali. E, come abbiamo visto, sono i controfattuali di cui abbiamo bisogno per risolvere i problemi dei free rider nello schema adattamentista. Se le teorie adattamentiste sono storiche anziché nomologiche, questo spiega perché i problemi dei free rider non si possono risolvere entro la cornice adattamentista. "È caduta 'perché è scivolata su una buccia di banana." È molto probabile che sia andata così; ma non c'è alcuna legge - nemmeno una legge statistica - che abbia "buccia di banana" nel suo antecedente e "scivola e cade" nel suo conseguente. Analogamente, Napoleone fu sconfitto a Waterloo perché pioveva da giorni e il terreno era troppo fangoso per la carica della cavalleria. (Così, perlomeno, ci dicono; e chi siamo noi per sostenere altrimenti?) Ma proprio non ne segue che esistano leggi che collegano la quantità di fango sul terreno con l'esito delle battaglie. I naturalisti metafisici (nelle cui file militiamo) supponiamo debbano dire che quello che è successo a Waterloo deve essere sussunto sotto qualche legge di copertura. Senza dubbio, per esempio, esemplifica (fra le altre cose) leggi della meccanica degli oggetti di medie dimensioni. Ma non ne segue che esistano leggi del fango così descritto, o delle battaglie così descritte, ancor meno delle connessioni causali fra di essi, così descritti; che è quel che sarebbe necessario se "fu sconfitto a causa del fango" deve essere un caso di un tipo di spiegazione per sussunzione sotto una legge o se deve sostenere dei controfattuali su quello che sarebbe successo se non avesse piovuto. Supponiamo, analogamente, che quando un t1 compete con un t2, qualche legge deve governare le interazioni causali fra di loro. La questione, però, è se si tratta di leggi della competizione, o se addirittura sono leggi della macrobiologia. Immaginiamo che Darwin non sarebbe contento se si scoprisse che, anche se c'è davvero una spiegazione della mutabilità delle specie, questa non sfrutta il vocabolario della competizione, della selezione e simili, bensì (e sarebbe possibile) il vocabolario della meccanica quantistica. Va di conserva con il fatto che non facciano ricorso a leggi di copertura, che le spiegazioni basate su narrazioni storiche sembrino spesso a posteriori. E il motivo per cui spesso sembrano tali è che di solito lo sono. Dato che sappiamo già chi ha vinto, possiamo raccontare una storia molto plausibile (del tipo "troppo fango sul terreno") sul perché non vinse Napoleone. Ma, non essendoci legge di copertura da citare, dubitiamo che Napoleone o Wellington o chiunque altro avrebbe potuto prevedere l'esito, prima dell'evento. La difficoltà è che ci sarebbe stata una storia plausibile per spiegare quello che era avvenuto, chiunque avesse vinto: previsione e retrovisione sono famose per questa asimmetria. Stando così le cose, ci sono moltissime spiegazioni storiche ragionevoli dello stesso evento, e non c'è nulla per scegliere l'una o l'altra. Wellington ha davvero vinto a causa del fango? O perché i reggimenti prussiani sono riusciti ad arrivare proprio per il rotto della cuffia? O semplicemente perché Napoleone aveva perso il suo tocco? (E intanto che siamo in tema di storia, che cosa, esattamente, ha causato la Riforma?) Non si discute del fatto che le competizioni fra individui con fenotipi diversi spesso abbiano esiti diversi; e ovviamente, in ogni caso, deve esserci qualche spiegazione del perché il vincitore ha vinto e il perdente no. Ma non c'è alcuna ragione per supporre che queste spiegazioni normalmente chiamino in causa leggi che si applicano agli individui in virtù dei loro tratti fenotipici. Stando così le cose, non deve per forza esserci. qualcosa da scegliere fra affermazioni relative ai controfattuali corrispondenti. I piccoli mammiferi hanno vinto la loro competizione con i grandi dinosauri. Ma ci sono riusciti grazie alla loro piccolezza? Dipende (fra le altre cose) dal fatto se avrebbero vinto anche se non ci fosse stato un meteorite. Possiamo raccontarvi una storia plausibile sul perché avrebbero potuto: gli animali di piccole dimensioni sono in grado di trafugare le uova di dinosauro, quando i dinosauri non vedono, per mangiarsele (il che è un male per la fitness dei dinosauri). D'altra parte, possiamo raccontarvi una storia plausibile del perché, se non ci fosse stato il meteorite, i mammiferi non avrebbero vinto: non ci sarebbe stata selezione per la tolleranza al cambiamento climatico, che i mammiferi avevano ma i dinosauri no. (Si noti che, secondo l'ultima storia, non c'è stata selezione per la piccolezza o la rapidità dei mammiferi, ma per la gamma di temperature che erano in grado di sopportare.) E allora, a quali controfattuali offrono sostegno le nostre narrazioni evoluzionistiche sull'estinzione dei dinosauri? A nessuno dei due? A entrambi? E, analogamente, per quale tratto l'evoluzione ha operato la selezione, quando ha selezionato individui che proteggevano i loro piccoli? Un interesse altruistico per la prole o un interesse egoistico per i propri geni? Be',...
C'è, tuttavia, un modello di spiegazione adattamentista
che sembra corrispondere piuttosto bene ai fatti. Se è altrimenti percorribile,
suggerisce che queste spiegazioni, anche
se non sono nomiche e non offrono sostegno ai controfattuali, hanno precedenti
perfettamente rispettabili. Se le spiegazioni adattamentiste sono specie di
narrazioni storiche, dal disastro si può salvare tutto tranne la selezione-per.
E va tutto bene, perché il meccanismo dell'evoluzione non è la selezione-per
tratti fenotipici. I meccanismi dell'evoluzione sono i
soggetti non della teoria evoluzionistica ma delle vignette che
la storia naturale ritaglia caso per caso. L'evoluzione è un tipo
di storia ed entrambe sono solo una dannata cosa dopo l'altra.
Conclusione retorica
Ecco un'analogia (in effetti, pensiamo, un'analogia molto stretta). Per ogni persona ricca, deve esserci qualcosa che spiega questa sua condizione: eredità, lasciti, cupidigia, acutezza, mendacità, oppressione dei poveri, essersi trovato al posto giusto nel momento giusto, avere amici in alto loco, pura e semplice fortuna, rapina al treno postale, quello che volete. Che cosa porti al diventare ricchi è, ovviamente, fortemente dipendente dal contesto: è a causa delle differenze di contesto che nessuno di noi oggi ha la minima probabilità di poter diventare ricco (per esempio) nel modo in cui lo fece Gengis Khan; o nel modo (non molto diverso) in cui lo diventò Andrew Carnegie; o nel modo (molto differente) in cui lo diventarono gli eredi di Andrew Carnegie; o nel modo (ancora molto diverso) in cui lo diventò Liberace; e così via. Analogamente, l'estrema sensibilità al contesto rende del tutto improbabile che ci possa essere una teoria del diventare ricchi in sé, nonostante tutti quei libri su come diventare ricchi che si vendono negli aeroporti. In particolare, è molto improbabile che ci siano generalizzazioni che sono leggi (quindi in grado di sostenere controfattuali, non ad hoc e non vuote, e così via) e che specificano le varie situazioni in cui è possibile diventare ricchi e le proprietà in virtù delle quali, se uno le avesse, diventerebbe ricco in quelle situazioni. Questo, notate bene, è pienamente compatibile con l'esistenza di storie completamente convincenti - storie da cui uno deve essere convinto - che spiegano, caso per caso, che cosa aveva un tizio, per cui è diventato ricco quanto lo è diventato nelle circostanze che vigevano quando e dove lo è diventato. Pensiamo che le spiegazioni adattamentiste dell'evoluzione dei tratti ereditabili siano davvero molto simili a queste. Quando funzionano è perché presentano delle narrazioni storiche plausibili, non perché citano delle leggi di copertura. In particolare, con buona pace dei darwinisti, l'adattamentismo non esplicita i meccanismi della selezione dei tratti fenotipici ereditabili; non potrebbe, perché non ci sono meccanismi della selezione dei tratti fenotipici ereditabili (in quanto tali). Tutto quello che c'è sono molti, molti modi differenti in cui vari individui riescono a prosperare nelle molte, molte situazioni ambientali in cui riescono a farlo. Diamond (in Mayr, 2001, p. x) nota che Darwin non ha semplicemente presentato "una ben elaborata teoria dell'evoluzione. Cosa ancor più importante, ha proposto anche una teoria della causazione, la teoria della selezione naturale". Bene, se abbiamo ragione, questo è proprio quello che Darwin non ha fatto: una "teoria della causazione" è esattamente quello che la teoria della selezione naturale non è. A ben pensarci, è esattamente quello che ancora non abbiamo. Dal punto di vista del filosofo della scienza, forse la conelusione di tutto questo è l'importanza di mantenere chiara la differenza fra spiegazioni storiche e spiegazioni nomologiche. Proprio come non c'è niente di ovviamente sbagliato nelle prime, non c'è niente di ovviamente sbagliato nelle seconde. Normalmente, iniziano con un mondo in cui sono specificate le condizioni iniziali e le leggi naturali, e deducono previsioni su quali situazioni si realizzeranno in quel mondo. È vero per definizione che la spiegazione di un evento per riferimento a una legge deve citare qualche proprietà dell'evento in virtù della quale la legge lo sussume. Niente ha una spiegazione nomologica a meno che non appartenga a un genere naturale. (Noi pensiamo che quello che abbiamo appena detto sia una serie di truismi.) Le spiegazioni nomologiche hanno goduto di buona pubblicità in filosofia, e giustamente. Che siano o meno i veri paradigmi della spiegazione scientifica, è del tutto chiaro di fatto che molte spiegazioni scientifiche sono leggi empiriche o incorporano un riferimento a leggi empiriche. Ma non c'è qualcosa di sbagliato neanche nelle spiegazioni costituite da narrazioni storiche. Sostanzialmente, una narrazione storica inizia con un evento per cui cerca di fornire una causa empiricamente sufficiente (il cavallo è andato perso per mancanza di un ferro). Perciò le narrazioni storiche sono intrinsecamente a posteriori (anche se non, ovviamente, intrinsecamente ad hoc). Le condizioni causalmente sufficienti che le narrazioni storiche chiamano in causa appartengono, in modi familiari, a catene di tali condizioni che (assumendo il determinismo) possono risalire indietro quanto si vuole (il ferro si staccò per la perdita di un chiodo, e così via). Fino a che punto debba risalire una simile spiegazione dipende, come si direbbe ridendo, da fattori pragmatici: che cosa si deve spiegare, a chi e a quale fine. Molte teorie scientifiche paradigmatiche sono, pensiamo, meglio intese come narrazioni storiche. Pensate, fra le altre, a teorie sulla geografia lunare, teorie sul motivo per cui i dinosauri si sono estinti, teorie sull'origine del Grand Canyon, o del sistema solare o, volendo guardare, dell'universo. Tutti questi progetti (e, sicuramente, molti altri) sono ricerche a posteriori di catene di condizioni causali sufficienti la cui soddisfazione spiegherebbe il verificarsi dell'evento in questione. Se abbiamo ragione, anche le teorie adattamentiste su come evolvono i tratti ereditabili sono di questo tipo. Questo solo per dire che una collezione dei vari meccanismi dell'adattamento non costituirebbe un genere naturale; perlomeno, non se il modello della spiegazione è la sussunzione sotto generalizzazioni nomologicamente necessarie. Ma se non ci sono generalizzazioni nomologicamente necessarie sui meccanismi dell'adattamento in quanto tale, allora la teoria della selezione naturale si riduce a una verità banale: "Se un tipo di individui prospera in un certo tipo di situazioni, allora deve esserci qualcosa in questi individui (o in queste situazioni, o in entrambi) in virtù di cui prospera". Beh, ovviamente deve esserci; persino un creazionista potrebbe essere d'accordo. Quel che rende così divertente leggere la letteratura adattamentista non sono le leggi dell'evoluzione che propone (non ne propone alcuna), e analogamente non sono i meccanismi del cambiamento fenotipico che scopre (non ne scopre alcuno). Sono le storie che racconta su quanti strani tipi di creature esistono; e su come, caso per caso, queste creature sono diventate così strane; e su come, strane come sono, ciascuna abbia in qualche modo trovato un sistema per sopravvivere. Ma, per ripetere la morale principale: dal fatto che esistano spiegazioni adattamentiste che spingono alla credenza razionale, non ne segue che esistano leggi dell'adattamento. E se non ci sono leggi dell'adattamento, non c'è modo (per quanto se ne sappia) di costruire una nozione di selezione-per che non sia semplicemente vuota. E la "selezione-per" non è una nozione di cui una spiegazione (neo)darwiniana dell'evoluzione possa fare a meno. | << | < | > | >> |Pagina 200Darwin era interessato a capire perché ci sono specie, cioè perché i fenotipi ricadono in famiglie, quando sono raggruppati per somiglianza. Pensava che le somiglianze fra cospecifici fossero in gran parte l'effetto della comunanza di antenati e, nonostante qualche controesempio, aveva certamente ragione su questo. Ma c'è una domanda diversa, che Darwin non si è posto, e la cui analisi avrebbe potuto dimostrarsi - in effetti potrebbe ancora dimostrarsi - altrettanto fruttuosa; e cioè perché certi fenomeni, perfettamente immaginabili, semplicemente non si danno, né qui e ora né in alcun punto della documentazione fossile. Perché, per esempio, non ci sono maiali con le ali? Sicuramente la risposta non sta in un ricorso alla selezione esogena per la fitness. Nessuno pensa che se non ci sono maiali con le ali è perché i maiali con le ali sono stati vittime di una selezione avversa nella competizione preistorica con i maiali senza ali. Piuttosto, i maiali non hanno ali perché sui maiali non c'è un posto dove metterle. Bisognerebbe cambiare in molti modi diversi un maiale, se gli si volessero aggiungere le ali. Bisognerebbe fare qualcosa al suo peso, alla sua forma, alla sua muscolatura, al suo sistema nervoso, e alle sue ossa; per non parlare del dotarlo di penne. Una volta che un organismo si trova sul percorso evolutivo che lo porta a diventare un maiale non c'è modo di aggiungere delle ali al suo fenotipo. Tutto questo, però, non ha un granché a che vedere con l'attività di forze esogene. Spesso, quello che spiega perché qualche fenotipo non si dia non è il fatto che la selezione abbia agito contro di esso, ma che certe combinazioni dei mattoncini fenotipici non sono possibili. Ci sono dei vincoli sui fenotipi che, per così dire, agiscono non dall'esterno ma dal basso.Quanti sono questi vincoli endogeni, e di che tipi sono? Nessuno lo sa esattamente. È chiaro, però, che quanti più sono i vincoli endogeni, tanto meno lavoro ha da svolgere l'ecologia per dare forma ai fenotipi. Al limite, non ha alcun lavoro da fare; al limite gli organismi sono come sono perché non potrebbero essere in altro modo. Se, a un certo stadio dell'evoluzione, gli unici fenotipi endogenamente possibili sono grandi dinosauri e piccoli mammiferi, allora la selezione non ha molto da fare per determinare il corso dell'evoluzione da quello stadio; tutto quello che serve è dare una botta in testa ai dinosauri con un meteorite.
Torneremo su questa e altre riflessioni analoghe nell'ultimo capitolo. Per
il momento basta questo:
- Non c'è bisogno di spiegare che gli organismi siano "raffinatamente adattati alle loro ecologie"; se è vero che lo sono, è una tautologia che lo siano. - Non c'è bisogno di spiegare che cosa c'è nel fenotipo di un organismo, grazie al quale è in grado di procurarsi i mezzi di sostentamento nell'ecologia che abita. Queste spiegazioni sono nel dominio di quella che si potrebbe chiamare "storia naturale sincronica"; le teorie dell'evoluzione non si prefiggono proprio di fornirle. Se si vuole capire in che cosa consista l'adattatività del fenotipo di un organismo, bisogna porsi la domanda (sincronica) "come fa a procurarsi i mezzi di sostentamento?", non la domanda (diacronica) "come è successo che ha potuto procurarsi i mezzi di sostentamento in questo modo?"
- Non c'è bisogno di spiegare come (cioè grazie a quale processo storico
effettivo) gli organismi sono arrivati ad avere i fenotipi che hanno. Questo è
il dominio di quella che si potrebbe chiamare "storia naturale diacronica", di
cui l'adattamentismo è una versione. Ma non c'è motivo ovvio
per supporre, come fanno i sostenitori della selezione naturale, che le
proprietà fenotipiche siano sostanzialmente
l'effetto dell'adattamento a variabili esogene.
Qualche volta si vince, qualche volta si perde: qualche volta la colpa è delle stelle, qualche volta è vostra, e qualche volta arriva in testa un meteorite. Sono cose che succedono, come osservava un famoso dinosauro. Di qui l'osservazione di Gould, che, se si facesse girare una seconda volta il nastro dell'evoluzione, molto probabilmente si otterrebbe qualcosa di diverso. La storia (compresa la storia naturale) riguarda quello che è successo effettivamente, non quello che doveva succedere, né quello che succederebbe se Madre Natura dovesse fare un altro tentativo. Quello che doveva succedere è il dominio della teoria, non della storia; e non c'è una teoria dell'evoluzione. | << | < | > | >> |Pagina 2029. Riepilogo e postludio"Va bene; se allora Darwin si sbagliava, quale pensate che sia il meccanismo dell'evoluzione?" Risposta breve: non sappiamo quale sia il meccanismo dell'evoluzione. Per quel che possiamo capire, nessuno sa esattamente come evolvono i fenotipi. Noi pensiamo che, molto probabilmente, evolvono in molti modi diversi; forse ci sono tanti tipi di percorsi causali distinti per la fissazione dei fenotipi quanti sono i tipi diversi di storie naturali degli organismi di cui sono i fenotipi (si veda il capitolo precedente). [...] [...] Quello che neghiamo, però, è che esista una teoria unitaria (per esempio una teoria unitaria delle interazioni organismo-ambiente) nei cui termini si spiegano questi fenomeni, tutti o la maggior parte di essi; o che i vari tipi di resoconti che li spiegano in generale comportino l'esistenza di leggi di selezione esogena. Forse non vi sembra un granché; forse preferireste che esistesse una teoria unificata - la selezione naturale - della fissazione evolutiva dei fenotipi. E così sia; ma noi possiamo sostenere qualcosa che i darwinisti non possono sostenere. Non c'è fantasma nella nostra macchina; né Dio, né Madre Natura, né Geni egoisti, né lo Spirito del Mondo, né intenzioni che volano libere; e neanche allevatori fantasma. Quel che nutre i fantasmi nel darwinismo è il suo ricorso nascosto a spiegazioni biologiche intensionali, di cui noi qui proponiamo di fare a meno. Darwin ha indicato la direzione per arrivare a una teoria pienamente naturalistica - in effetti pienamente ateistica - della formazione dei fenotipi; ma non ha visto come arrivarci fino in fondo. Ha eliminato Dio, se volete, ma Madre Natura e altri pseudo-agenti ne sono usciti indenni. Pensiamo che sia ora di liberarci anche di loro. | << | < | > | >> |Pagina 233Un dialogo amichevole con Telmo PievaniSono grato a Telmo Pievani , professore di Filosofia della scienza presso l'Università degli Studi di Milano-Bicocca, qualificatissimo teorico dell'evoluzione, profondo conoscitore dell'intera opera darwiniana, formatosi anche negli Stati Uniti con due dei migliori evoluzionisti non strettamente darwinisti ( Niles Eldredge e Ian Tattersall ), in sintonia con i lavori di Stephen Jay Gould e Richard Lewontin. Come alludevo qui sopra, molti punti ci accomunano e alcuni invece ci dividono. Mi dice che ritiene giustissimo criticare la tracimazione del neo-darwinismo stretto sulle scienze umane in genere e sulla psicologia in particolare. Avrebbe consigliato di imperniare tutto questo libro su questa critica, piuttosto che relegarla in un'appendice. Condivide il giudizio sull'importanza di tutto quanto esponiamo nella Parte prima, sottolineando, però, che si tratta di scoperte tra loro piuttosto, per adesso, eterogenee. Ne convengo e penso che questo emerga proprio da come le presentiamo. Sottolinea anche che il ritorno delle leggi della forma non morde ancora dentro la ricerca evoluzionistica giorno per giorno e che i fattori fisici, chimici e auto-organizzativi sono lungi dall'esaurire i meccanismi dell'evoluzione. Per ora resta, infatti, spesso arduo tradurli in spiegazioni specifiche e in predizioni dettagliate. Di nuovo siamo d'accordo e penso proprio che lo diciamo espressamente. Vengo ora a ciò che ci divide. Come anche il genetista e alto divulgatore Edoardo Boncinelli , Massimo Pigliucci, Eldredge, Tattersall, Eva Jablonka e altri darwiniani dichiaratamente eterodossi, Pievani considera tutto ciò un allargamento della teoria della selezione naturale, un pluralismo darwiniano che va propagato e rafforzato, ma non un sovvertimento del neo-darwinismo, come invece Fodor e io sosteniamo. In un suo lungo messaggio mi dice, tra l'altro: Non c'è quindi un'intera "nuova biologia" emergente, ma una evoluzione (sì profonda) del programma di ricerca neodarwiniano; altrimenti le citazioni che usate, estrapolate dal contesto, sembrano assumere un significato troppo radicale, che non hanno nella realtà (come nel caso di mutazioni sistemiche improvvise che darebbero origine, normalmente, a nuove specie: non è così se non in casi circoscritti). Secondo me questo è fuorviante rispetto al reale dibattito in corso: credo che avessero proprio ragione Gould e Lewontin nel 1979 a prevedere che la selezione naturale sarebbe stata ampiamente riconsiderata nel suo funzionamento come fattore non unico di cambiamento evolutivo, ma pur sempre "il più importante". Qui, in effetti, dissentiamo. Non è più sostenibile che la selezione naturale sia il fattore più importante del cambiamento evolutivo, come abbiamo ampiamente visto in questo libro. L'onere della prova ricadeva, fino a ieri, su chi sosteneva non lo fosse, ma oggi ricade su chi sostiene che lo sia. Ed è un onere che diventa sempre più insostenibile. Quanto a un "allargamento" (o un'estensione, secondo la dizione propugnata insistentemente da Massimo Pigliucci) del neo-darwinismo, posso solo controbattere che le citazioni di biologi sperimentali riportate nel libro e quelle ora riportate in questo aggiornamento per l'edizione italiana propendono verso una svolta radicale, non verso un allargamento. E sono in numero crescente i biologi che la pensano così, quasi di mese in mese. Pur con tutto l'apprezzamento per i lavori di Boncinelli, Pigliucci, Eldredge, Tattersall e dello stesso Pievani, noi ci sentiamo più vicini ai Gibson, Margulis , Ayala , Koonin, Newman e Kruglyak di questo (nuovo) mondo. Può darsi che si tratti solo di una differenza di interpretazione, o perfino di definizione ("di quale darwinismo si parla?", raccomanda Pievani di precisare sempre). O può invece darsi che, quando questo "allargamento" continuerà a estendersi ulteriormente, come è prevedibile che farà, perfino il neo-darwinismo eterodosso e poliedrico si troverà spiazzato oltre misura e, forse senza ammetterlo, si avvicinerà a queste posizioni non conciliatorie. Infine, Pievani ci tira garbatamente l'orecchio per aver contrapposto le spiegazioni eterogenee della storia naturale a quelle legi-simili (nomologiche, cioè desunte da una legge generale) della teoria della selezione naturale. Riporto le sue stesse parole: La pervasività esplicativa della selezione naturale non è compromessa dal suo operare in un contesto di vincoli fisici e strutturali, né dal suo operare storico, imperfetto, contingente (già ampiamente presente in Darwin); qui secondo me davvero vi affidate troppo alla radicalizzazione retorica anti-Dawkins del vostro argomento, ma così date troppa importanza a chi ha trasformato Darwin in una formuletta divulgativa.
Rispondo con una domanda. Prendiamo una delle grandi
teorie della storia propugnate nel passato (sia essa idealista,
marxista, per cicli, per stadi o per sfida-e-risposta alla Toynbee o quant'altro
si vuole). Se ogni spiegazione di un evento
storico concreto poi richiede una massiccia e inevitabile aggiunta di dati
contingenti e di processi idiosincratici, possiamo dire che quella teoria è
valida? Che è davvero una teoria?
|