|
|
|
|
|
|
|
| << | < | > | >> |Indice
Perché solo noi
9 1. Perché adesso?
L'evoluzione della teoria evoluzionistica, 22
Modello tripartito, apprendimento vocale e genomica, 44
57 2. L'evoluzione della biolinguistica
89 3. L'architettura del linguaggio e la sua importanza per l'evoluzione
106 4. Triangoli nel cervello
Fuori della portata della selezione naturale?, 106
Che cosa?, 108
Chi?, 135
Dove e quando?, 144
Come?, 151
Perché?, 155
159 Bibliografia
179 Ringraziamenti
181 Indice analitico
|
| << | < | > | >> |Pagina 91.
Perché adesso?
Siamo nati gridando, ma quelle grida annunciano le prime stimolazioni del linguaggio. Le grida dei bambini tedeschi riflettono la melodia della lingua tedesca; quelle dei bambini francesi riflettono la lingua francese, a quanto pare acquisita in utero (Mampe e al. 2009). Entro il primo anno circa di vita i neonati padroneggiano il sistema fonico della loro lingua; poi, trascorsi alcuni anni, entrano in conversazione con coloro che li accudiscono. Questa straordinaria capacità specie-specifica di acquisire qualsiasi lingua umana (la «facoltà del linguaggio») ha da tempo sollevato importanti questioni biologiche, incluse le seguenti: qual è la natura del linguaggio? come funziona? come si è evoluto? La presente raccolta di saggi affronta la terza questione: l'evoluzione del linguaggio. Nonostante le affermazioni contrarie, in verità è sempre esistito un forte interesse per l'evoluzione del linguaggio sin dagli albori della grammatica generativa a metà del Novecento. La grammatica generativa cercava, per la prima volta, di fornire resoconti espliciti delle lingue (delle grammatiche) che spiegassero quella che chiamiamo «proprietà fondamentale del linguaggio», secondo cui una lingua è un sistema computazionale finito che produce un'infinità di espressioni, ciascuna delle quali ha un'interpretazione ben precisa nel sistema semantico-pragmatico e nel sistema sensomotorio (detto alla buona: pensiero e suono). Quando questo problema fu proposto per la prima volta, il compito sembrava soverchiante. I linguisti si affannavano a costruire grammatiche a malapena adeguate e i risultati erano così complessi che all'epoca era chiaro che non si sarebbero potuti evolvere. Per questo motivo le discussioni sull'evoluzione del linguaggio raramente arrivavano alla pubblicazione, sebbene ci fossero alcune notevoli eccezioni. E allora che cosa è cambiato? Tanto per cominciare, la teoria linguistica è maturata. Adesso i complessi sistemi di regole linguistiche appartengono al passato: sono stati sostituiti da impostazioni molto più semplici e quindi più plausibili da un punto di vista evolutivo. Inoltre anche certi componenti biologici fondamentali associati al linguaggio, in particolare il sistema input-output dell'apprendimento e della produzione vocale, che fa parte del sistema che chiameremo «esternalizzazione», sono stati chiariti biologicamente e geneticamente, tant'è che possiamo impiegare effettivamente una strategia divide et impera e mettere da parte l'aspetto sensomotorio dell'esternalizzazione mentre focalizziamo proprietà del linguaggio più centrali. Sebbene molto debba rimanere incerto semplicemente perché non disponiamo delle prove necessarie, gli sviluppi della teoria linguistica nel corso delle ultime due decadi hanno notevolmente chiarito certi aspetti dell'origine del linguaggio. In particolare, oggi abbiamo buone ragioni per ritenere che un componente essenziale del linguaggio umano (il motore principale che aziona la sintassi del linguaggio) sia molto più semplice di quanto molti avessero pensato pochi decenni or sono. Si tratta di un risultato ben accolto sia dalla biologia evoluzionistica sia dalla linguistica. I biologi sanno bene che quanto più delimitata è la definizione del «fenotipo» (alla lettera: la «forma che appare» esternamente) tanto migliore è la nostra padronanza biologica del modo in cui il fenotipo si sarebbe evoluto. E analogamente: tanto più stretto è il divario tra noi e le specie che sono prive del linguaggio. Avendo a disposizione questo fenotipo meglio definito, possiamo iniziare a risolvere il dilemma che afflisse fin dall'inizio la spiegazione darwiniana dell'evoluzione del linguaggio. [...] Contrariamente a quanto taluni credono, la discussione sull'evoluzione del linguaggio come «problema di Darwin» non fu un argomento tabù fino alla sua «rinascita» negli anni novanta, come qualche parente eccentrico relegato per trent'anni in soffitta. Al contrario, era un argomento di profondo interesse a Cambridge (Mass.) nel corso degli anni cinquanta e sessanta, e poi per tutti gli anni settanta. Questo profondo interesse si riflette direttamente nella prefazione che Eric Lenneberg scrisse nel settembre 1966 a Biological Foundations of Language, dove ricorda il suo debito «durante i quindici anni trascorsi a Cambridge» (1967, trad. it. p. 3) nei confronti di un elenco di nomi famosi e ben noti: Roger Brown , Jerome Bruner , George Miller , Hans-Lukas Teuber, Philip Liberman, Ernst Mayr , Charles Gross, e anche Noam Chomsky. Secondo noi, il libro di Lenneberg rimane ancora estremamente pertinente: in particolare il capitolo 6, «Il linguaggio alla luce dell'evoluzione e della genetica», resta ancora un modello di pensiero evolutivo sfaccettato, com'è anche un suo lavoro precedente (Lenneberg 1964). In un certo senso i nostri saggi aggiornano ciò che Lenneberg aveva già scritto. Se capiamo bene questa storia, fu Lenneberg a proporre con preveggenza una collezione longitudinale di linguaggi orientati ai bambini; scoprì l'invenzione spontanea del linguaggio dei segni come linguaggio umano completo (alla Perkins School per non udenti, Watertown, Mass.); trovò che l'acquisizione del linguaggio riesce ancora nonostante importanti patologie; presentò le prove a favore di un periodo critico per l'acquisizione del linguaggio; rilevò certe dissociazioni tra il linguaggio sintattico e altre facoltà cognitive; coniò la terminologia moderna, per esempio il «cervello pronto per il linguaggio»; utilizzò l'analisi genealogica delle famiglie con menomazioni linguistiche, richiamando i dati concernenti il gene FOXP2 per fornire le prove che il linguaggio ha un componente genetico; e osservò che «non vi è alcuna necessità di ammettere che esistano dei "geni del linguaggio"» (Lenneberg 1967, trad. it. p. 298). Mise anche in contrasto le impostazioni continuistiche con quelle discontinuistiche nello studio dell'evoluzione del linguaggio, argomentando a favore della posizione discontinuistica, convalidata in parte da prove fondamentali quale l'evidente uniformità della facoltà linguistica: «L'esistenza di capacità linguistiche identiche per tutte le razze fa pensare che questo fenomeno debba essere comparso prima della diversificazione razziale» (ibid.). In realtà è sempre esistito un interesse costante per la questione del linguaggio e della sua evoluzione. Certo, negli anni cinquanta e sessanta non si sarebbe potuto dire sull'evoluzione del linguaggio molto più di quanto scritto da Lenneberg. Le grammatiche generative tipiche dell'epoca consistevano in molte regole trasformazionali, complesse e ordinate. Un'occhiata all'appendice II delle Syntactic Structures di Chomsky (1957), con le sue ventisei regole estremamente particolareggiate per un frammento della lingua inglese, rivela subito questa complessità. | << | < | > | >> |Pagina 17Ricapitoliamo allora quello che ci dice la nostra analisi minimalista. Nel migliore dei casi rimane una singola operazione per costruire la struttura gerarchica richiesta dalla sintassi del linguaggio umano: Fondere. Questa operazione prende due elementi sintattici qualsasi e li combina in un'espressione nuova, più estesa e strutturata gerarchicamente.Nei suoi termini più semplici, l'operazione Fondere è esattamente la formazione di un insieme. Dato un oggetto sintattico X (o un atomo somigliante a una parola o qualcosa che a sua volta è un prodotto di Fondere) e un altro oggetto sintattico Y, Fondere forma un nuovo oggetto gerarchicamente strutturato come insieme {X, Y}; al nuovo oggetto sintattico si assegna anche un'etichetta mediante un algoritmo che soddisfa la condizione della computazione minima. Per esempio, dati read e books, Fondere li combina in {read, books} e il risultato è etichettato attraverso una ricerca minima che localizza gli aspetti della «testa» della combinazione in questo caso gli aspetti dell'elemento verbale read. Tutto ciò concorda con l'idea tradizionale, secondo la quale la struttura a costituenti di read books è un «sintagma verbale». Questa nuova espressione sintattica può poi entrare in altre computazioni, cogliendo quella che in precedenza abbiamo chiamato «proprietà fondamentale» del linguaggio umano. Particolari ulteriori su questa impostazione si possono trovare negli altri capitoli, ma per il momento sia chiaro che circoscrivendo in maniera limitata il fenotipo si allegerisce notevolmente l'onere esplicativo della teoria evoluzionistica: semplicemente non ci resta molto da spiegare, riducendo il paradosso darwiniano. Il perfezionamento e la delimitazione recenti del fenotipo del linguaggio umano sono la prima motivazione alla base di questa raccolta di saggi. La seconda motivazione è il miglioramento della nostra comprensione del fondamento biologico del linguaggio. Adesso possiamo impiegare effettivamente una strategia divide et impera per scomporre il difficile problema evolutivo del «linguaggio» nelle tre parti definite dalla Proprietà Fondamentale: 1) un sistema computazionale interno che costruisce espressioni strutturate gerarchicamente con interpretazioni sistematiche all'interfaccia con due altri sistemi interni, ossia: 2) un sistema sensomotorio per l'esternalizzazione come produzione o analisi sintattica e 3) un sistema concettuale per l'inferenza, l'intepretazione, la programmazione e l'organizzazione dell'azione, quello che informalmente si chiama «pensiero». È importante notare che l'esternalizzazione include molto più del semplice apprendimento e produzione vocali/motorii, comprendendo perlomeno certi aspetti del linguaggio quali la formazione delle parole (morfologia) e la sua relazione con i sistemi fonici del linguaggio (fonologia e fonetica), il riassestamento in uscita, per alleggerire il carico mnemonico durante la produzione, e la prosodia. Più importante, tuttavia, dal nostro punto di vista è a quanto pare la possibilità, nel caso del linguaggio, di usare qualunque modalità sensoriale per l'ingresso o l'uscita: suono, segno o tatto (per fortuna l'olfatto risulta assente da questo elenco). Si noti che la struttura gerarchica interna non reca in sé alcuna informazione circa l'ordine da sinistra a destra dei sintagmi, delle parole o di altri elementi. Per esempio, le possibilità verbo-oggetto o oggetto-verbo, che distinguono il giapponese dall'inglese e dal francese, non sono nemmeno rappresentate nella struttura gerarchica interna. Invece l'ordine temporale sequenziale del linguaggio è imposto dalle esigenze dell'esternalizzazione. Se la modalità è uditiva, questa uscita si chiama più familiarmente «discorso» e include l'appredimento e la produzione vocale. Ma la modalità in uscita può anche essere visiva e motoria, come nei linguaggi dei segni. Grazie in parte agli studi comparativi, neurofisiologici e genomici sugli uccelli canori, il fondamento biologico dell'apprendimento vocale sta per essere compreso come sistema evolutivamente convergente: un sistema che si è evoluto negli uccelli e in noi in maniera identica, ma indipendente. [...] Passando ora alla nostra terza motivazione, ci è sembrato che le importanti idee di Lenneberg sulla biologia e la natura dell'evoluzione linguistica corressero il pericolo di andare perdute. Per esempio, egli aveva discusso attentamente le ragioni a favore e a sfavore delle impostazioni basate sulla «continuità» evolutiva, come quella di Darwin, in opposizione a quelle basate sulla «discontinuità», che aveva preferito. La scelta era apparsa particolarmente acuta, dati i recenti progressi del pensiero evoluzionistico che hanno chiarito queste posizioni. Come qualsiasi campo scientifico fertile, la biologia evoluzionistica moderna si è allontanata dall'originaria concezione darwiniana dell'evoluzione come cambiamento adattivo risultante dalla selezione tra individui. Effettivamente Darwin si fece idee sbagliate su alcune cose. Forse è più noto ciò a cui rimediò la cosiddetta Sintesi Moderna: il matrimonio celebrato a metà del XX secolo tra l'evoluzione per selezione naturale e il mendelismo, in particolare i fattori ereditari (geni), che pose rimedio alla mancanza in Darwin di un buon modello di ereditarietà e che alla fine condusse alla moderna era genomica nell'analisi evoluzionistica. Darwin aveva adottato la teoria (sbagliata) dell'ereditarietà in voga ai suoi giorni: «l'ereditarietà mista». In base ai riscontri sulla mescolanza, se incrociamo fiori rossi con fiori bianchi, tutti i colori dei fiori risultanti cadranno nel mezzo: rosa. La mescolanza elimina rapidamente la variazione che la selezione naturale alimenta. Ricordiamoci di quando, da bambini, prendevamo un pennello bagnato e lo facevamo scorrere su una tavolozza di acquerelli: lo spettro dei singoli colori dal porpora al giallo si trasforma in un marrone opaco. Ma se tutta la stirpe ha gli stessi caratteri marrone opaco, la selezione naturale non ha più nulla da scegliere. Nessuno è al di sopra della media e nessuno è al di sotto della media; tutti sono uguali nel setaccio della selezione naturale. Se non c'è variazione, non c'è selezione naturale, e il meccanismo darwiniano si blocca in una o due generazioni. Ciò che occorre è un qualche modo per preservare la variazione da una generazione all'altra, anche se gli incroci tra fiori rossi e bianchi talvolta risultano rosa. Fu Mendel a scoprire la risposta: l'ereditarietà lavora attraverso particelle discrete (i geni), anche se naturalmente all'epoca egli non aveva modo di saperlo. Nella prima metà del XX secolo toccò ai fondatori della Sintesi Moderna (Sewall Wright, Ronald A. Fisher e J. B. S. Haldane ) mostrare come combinare in maniera sistematica l'ereditarietà particellare di Mendel con l'evoluzione darwiniana per selezione naturale, costruendo modelli matematici che dimostravano esplicitamente in che modo la macchina darwiniana potesse funzionare di generazione in generazione per modificare la frequenza dei caratteri nelle popolazioni. Ma Darwin ebbe anche il grave torto di presumere (in genere, implicitamente) che le popolazioni biologiche siano infinite, e che perfino nelle popolazioni realmente infinite l'evoluzione per selezione naturale sia un processo puramente deterministico. Ogni ingranaggio nella macchina dell'evoluzione (adattamento, migrazione, fertilità, accoppiamento, sviluppo ecc.) è soggetto ai colpi di fionda e ai dardi dell'oltraggiosa sorte biologica. Molto spesso la sopravvivenza del più adatto si riduce alla sopravvivenza del più fortunato, e questa circostanza incide sulla possibilità che l'evoluzione sia un processo uniformemente continuo come Darwin aveva immaginato. Per rendercene conto occorre un'analisi matematica più sottile e, per quanto possiamo vedere, nessuno dei libri recenti sull'evoluzione del linguaggio sembra averlo capito fino in fondo. Darwin stesso così osservava nella sua autobiografia: «La mia capacità di seguire un lungo e puramente astratto filo di pensiero è molto limitata; e perciò non me la sono mai cavata con la metafisica o la matematica» (Darwin 1887, p. 140). Nella parte restante del capitolo spacchettiamo le due ultime motivazioni in ordine inverso, cominciando con la teoria evoluzionistica e facendo seguito con uno sguardo all'impostazione divide et impera, unitamente all'evoluzione e alla genomica. Riserviamo ai capitoli 2 e 3 ulteriori particolari sul programma minimalista e la tesi minimalista in senso forte. | << | < | > | >> |Pagina 22L'evoluzione della teoria evoluzionisticaPer cominciare: che cosa c'è di tanto diverso nella teoria evoluzionistica contemporanea e nelle teorie sull'evoluzione del linguaggio? Possiamo partire con lo scenario storico intorno al 1930, l'apogeo della Sintesi Moderna, esposta sopra. La maggior parte di coloro che hanno scritto sull'evoluzione del linguaggio sembra aver compreso la storia del problema darwiniano dell'ereditarietà, unitamente alla sua soluzione attraverso la Sintesi Moderna. Alcuni fanno anche notare certi effetti semplici dell'ampiezza finita della popolazione sul cambiamento evolutivo: per esempio, gli effetti di campionamento in piccole popolazioni, talvolta chiamati «deriva genetica», possono condurre alla perdita di caratteri vantaggiosi indotta dalla cattiva sorte (la loro frequenza si riduce a zero nella popolazione) o alla fissazione completa di caratteri non vantaggiosi indotta dalla buona sorte (la loro frequenza sale a 1). Non è difficile capire perché. [...] Tuttavia, nella misura in cui siamo stati in grado di stabilirlo, nonostante l'adozione della Sintesi Moderna da parte di autori contemporanei, nessuno dei recenti resoconti sull'evoluzione del linguaggio umano sembra aver compreso appieno lo spostamento dal darwinismo convenzionale alla sua versione moderna interamente stocastica, in particolare il fatto che esistono effetti stocastici causati non soltanto da un campionamento quale la deriva non direzionale, ma anche dalla variazione stocastica diretta nell'adattamento, nella migrazione e nell'ereditarietà - anzi, tutte le «forze» che influiscono sulle frequenze individuali o genetiche. L'adattamento non è un «acido algoritmico universale» onnipotente, come alcuni hanno ritenuto. Le circostanze e il caso svolgono un ruolo importante. L'ambito delle possibilità è talmente vasto che molte «soluzioni», se non la maggior parte, sono irraggiungibili dall'evoluzione per selezione naturale, nonostante gli eoni di tempo e i bilioni di organismi a sua disposizione. Lungo queste direttrici alcuni risultati formali sono stati raggiunti di recente da Krishendu Chatterjee e colleghi (2014), i quali dimostrano che in genere il tempo richiesto dall'adattamento sarà esponenziale nella lunghezza della sequenza genomica: vale a dire, non c'è abbastanza tempo, anche dati gli eoni geologici. (La «capacità di elaborazione in parallelo», talvolta attribuita all'evoluzione per selezione naturale perché sono in gioco parecchi organismi, risulta essere una chimera). Illustriamo un effetto stocastico con un esempio tratto dal mondo reale. [...] Qual è la conclusione? Dobbiamo iniettare biologia del mondo reale e comportamento stocastico nel quadro evoluzionistico. Ciò comprende tassi di migrazione stocastici ( Ellis Island ieri e oggi), modelli stocastici dei fattori ereditari (dopotutto tu non assomigli ai tuoi nonni), interazioni tra geni (non esiste un singolo «gene del linguaggio») e fluttuazioni dell'adattamento ogni volta che la frequenza aumenta (sovrappopolazione in qualunque caso?). Se facciamo tutto questo, allora la concezione semplicistica, secondo la quale l'evoluzione adattiva raggiunge inesorabilmente picchi di adattamento, va in frantumi. È difficile «soddisfare» simultaneamente gli effetti di mille e più geni interagenti, per non parlare di sintonizzarli congiuntamente all'adattamento ottimale. | << | < | > | >> |Pagina 30Lo slittamento dal darwinismo deterministico alla sua versione pienamente stocastica è il risultato di una più sofisticata comprensione matematica e biologica dell'evoluzione e dei processi stocastici rispetto all'epoca in cui apparve On the Origin of Species (Darwin 1859). Questo progresso è prevedibile in ogni campo scientifico fiorente (l'evoluzione della teoria evoluzionistica stessa), ma a quanto pare molti studiosi hanno continuato a non deflettere dall'originaria idea darwiniana dell'evoluzione come selezione degli individui esclusivamente adattiva. Da tempo sappiamo, grazie a ricerche sia teoriche sia empiriche, che le concezioni di Darwin e della Sintesi Moderna non sempre sono accurate, ed esistono abbondanti prove sul campo per sostenerlo (Kimura 1983, Orr 1998 e 2005a, Grant e Grant 2014, Thompson 2013), e questo senza dover rifiutare in blocco il darwinismo, appellarsi alla trasmissione virale, al flusso genico orizzontale su vasta scala o alle macromutazioni miracolose, o incorporare legittime intuizioni dal campo dell'evoluzione e dello sviluppo, o «evo-devo».E allora, come si evolvono gli organismi? Si tratta di evoluzione per lento avanzamento o di evoluzione per sobbalzi, come fu detto nel famoso dibattito tra Stephen J. Gould e i suoi critici? (Turner 1984, Gould 2007). Entrambe le cose, ovviamente. Talvolta il cambiamento evolutivo adattivo è davvero molto lento e faticoso, in azione per milioni di anni secondo l'idea classica di Darwin. Ma talvolta il cambiamento evolutivo, e anche i cambiamenti comportamentali su larga scala, quali le preferenze alimentari del macaone (Thompson 2013, p. 65), possono essere relativamente rapidi, perfino mozzafiato. Una velocità che è stata confermata in centinaia di specie differenti attraverso tutti i maggiori gruppi filogenetici, come è stato fatto notare recentemente in una rassegna magistrale di John Thompson (ibid.). Qui non si devono confondere le acque ammettendo semplicemente, come fanno alcuni, che il gradualismo infinitesimale darwiniano talvolta affretti il passo. Siamo d'accordo. Ma la questione cruciale è il ritmo che concerne le prossime innovazioni evolutive. La nostra concezione abbraccia sia le possibilità a lunga scadenza, ossia milioni di anni e centinaia di migliaia di generazioni, come, a quanto pare, nell'evoluzione di un apparato di apprendimento vocale antecedente agli uccelli e a noi; sia le possibilità a breve termine, ossia alcune migliaia di anni e alcune centinaia o un migliaio di generazioni, come nel caso di adattamenti relativamente recenti quali la capacità dei Tibetani di crescere vigorosi alle alte altitudini dove c'è meno ossigeno, o la capacità di digerire il lattosio dopo l'infanzia nelle culture agricole casearie (Bersaglieri e al. 2004), o, secondo la nostra tesi centrale, la capacità innovativa di assemblare gerarchicamente la struttura sintattica. [...] Dunque possono effettivamente esserci stati slittamenti genomici/fenotipici repentini, che «spostano il punto di partenza da cui agisce la selezione», come afferma il biologo Nick Lane (2015, p. 312). Qui Lane fa notare lo spostamento straordinario e apparentemente unico e repentino dalla semplice vita cellulare, quella dei procarioti (con DNA circolare, privi di nucleo, asessuati e sostanzialmente senza morte biologica), alla gastronomia che condusse alla vita complessa, agli eucarioti, noi compresi (con DNA lineare, mitocondri, un nucleo, organelli complessi e, infine, al di là di Woody Allen, sesso, amore, morte e linguaggio). Come Lane fa notare, «non si deve confondere il salto genetico con l'adattamento» (ibid., p. 313). Dalla prospettiva del tempo geologico, questi cambiamenti furono rapidi. Tutto ciò evidenzia il ruolo del caso, della contingenza e del contesto biochimico-fisico nel cambiamento evolutivo innovativo: l'evoluzione per selezione naturale lavora alla cieca, senza avere in mente il «fine» di un'intelligenza superiore o il linguaggio. Alcuni eventi accadono soltanto una volta e non sembra che siano facilmente ripetibili: l'origine delle cellule con nuclei e mitocondri, e poi il sesso ecc. Altri biologi evoluzionisti sono d'accordo. Ernst Mayr , in un ben noto dibattito con Carl Sagan , faceva osservare che anche la nostra intelligenza - e quindi il linguaggio - probabilmente rientra nella stessa categoria: Nulla dimostra l'improbabilità dell'origine dell'intelligenza superiore meglio dei milioni di stirpi che non riuscirono a raggiungerla. Ci sono stati miliardi, forse addirittura cinquanta miliardi di specie fin dall'origine della vita. Soltanto una di queste ha raggiunto il tipo di intelligenza necessario per fondare una civiltà ... Riesco a immaginare soltanto due ragioni possibili di questa eccezionalità. Una è il fatto che l'intelligenza superiore non è affatto favorita dalla selezione naturale, contrariamente a quanto ci aspetteremmo. In effetti, tutti gli altri tipi di organismi viventi, milioni di specie, se la cavano bene senza un'intelligenza superiore. L'altra ragione possibile dell'eccezionalità dell'intelligenza è la straordinaria difficoltà di acquisirla ... non a caso perché i cervelli hanno un fabbisogno di energia estremamente elevato ... un cervello di grandi dimensioni, che consenta un'intelligenza superiore, si è sviluppato in meno dell'ultimo 6% della vita sulla linea degli ominidi. A quanto pare, ci vuole una combinazione complessa di rare circostanze favorevoli per produrre un'intelligenza superiore. (Mayr 1995) | << | < | > | >> |Pagina 36Riflettendovi, lo stesso dilemma darwiniano sorge con ogni novità genuina. Nel caso dell'origine dell'occhio Walter Gehring (2011) ha proceduto a un'analisi più sottile. L'occhio è il prodotto sia del caso sia della necessità, proprio come Jacques Monod (1970) aveva anticipato. Per il prototipo dell'occhio sono richieste due componenti: la cellula fotorecettore e la cellula pigmentata. La formazione iniziale del fotorecettore fu un evento casuale; non avvenne per selezione in seguito a una laboriosa ricerca incrementale «prova ed errore»: la cattura delle molecole di pigmento sensibile alla luce da parte delle cellule, regolata successivamente dal gene Pac-6. Ciò che un osservatore vedrebbe dall'esterno è un lunghissimo periodo di tempo geologico in cui la vita non aveva un pigmento di cellule fotorecettive, e poi la comparsa relativamente rapida di cellule-più-pigmento: o il pigmento fu catturato o non lo fu. Tutto ciò avvenne senza che fossero necessarie «numerose» e «lievi modificazioni». Certo, la molecola dovette passare per il setaccio della selezione e da allora è stata messa a punto, però dopo l'evento critico. Analogamente, la cellula pigmentata prototipica sorse dall'onnipresente pigmento melanina presente in una singola cellula insieme con il pigmento fotorecettivo ormai catturato. Poi, a un certo punto, ecco la singola cellula scindersi in due: di nuovo un evento stocastico, evidentemente sotto il controllo di un gene regolatore per differenziare le cellule. Anche in questo caso, se lo si osservasse «dall'esterno», si vedrebbe un periodo di stasi relativamente lungo, seguito dalla scissione tutto-o-nulla in due cellule: le figlie furono prodotte o non lo furono. «Da queste considerazioni concludiamo che il prototipo darwiniano dell'occhio sorse da una singola cellula per differenziazione cellulare, dove Pax6 controllava la cellula fotorecettore e Mitf controllava la cellula pigmentata» (Gehring 2011, p. 1058).| << | < | > | >> |Pagina 41Quale lezione trarre da questo accoglimento moderno del darvinismo e del cambiamento evolutivo? Fondamentalmente si ottiene ciò che si paga, e se lo si paga, si dovrebbe capire ciò che si è comprato: l'intero pacchetto con tutte le sue conseguenze. Se optiamo per il modello di Fisher, allora abbracciamo necessariamente il micromutazionismo, e già abbiamo escluso per decreto tutto quanto, tranne la selezione naturale, come driver causale dell'evoluzione del linguaggio. Come abbiamo visto, non siamo più in grado di spiegare l'origine di cellule complesse da cellule semplici procariote, l'origine degli occhi e molto altro ancora. D'altro canto, se non compriamo il modello di Fisher e passiamo alla concezione più moderna, lasciamo la porta aperta a un insieme più ricco di possibilità.Tornando alla storia umana, un esame dei reperti paleoarcheologici del nostro lignaggio Homo conferma il quadro non gradualistico, non quello gradualistico: una configurazione ricorrente di «sconnessioni tra epoche di apparizione (e scomparsa) di nuove tecniche e nuove specie» (Tattersall 2008, p. 108). Il punto fondamentale si capisce facilmente. Secondo Ian Tattersall , ogniqualvolta è comparsa una nuova specie Homo morfologicamente distinta, non si sono avute simultanee innovazioni tecniche o culturali. Invece le innovazioni tecnico-culturali compaiono molto dopo la comparsa di ogni nuova specie Homo, con un tempo misurato in centinaia di migliaia di anni. In altre parole, come scrive Tattersall (ibid., p. 103), «le innovazioni tecniche non sono associate con l'emergere di nuovi tipi di ominidi». [...] In ogni caso, il risultato dell'esodo dall'Africa dei nostri antenati fu che quella particolare specie Homo noi alla fine sarebbe riuscita a dominare il mondo, avrebbe assorbito quanto di buono ci fosse nei genomi di Neanderthal e Denisova, e avrebbe lasciato il resto: forse un quadro fantasioso, ma fin troppo familiare e nondimento inquietante, dato quel che sappiamo dalla storia successiva della nostra specie.
Quel che proprio non vediamo è un qualsiasi tipo di «gradualismo» nelle
nuove tecniche degli utensili, o in innovazioni quali il
fuoco, i ricetti o l'arte figurativa. L'uso controllato del fuoco compare
approssimativamente un milione di anni fa, un bel mezzo milione di anni dopo
l'emergere di
Homo ergaster.
Tattersall fa notare che il tipico
pattern
di stasi seguita da salti innovativi è compatibile con la nozione di
exaptation,
cioè l'evoluzione per selezione naturale coopta sempre caratteri esistenti per
usi nuovi; non può darsi «preveggenza» circa l'utilità futura di un carattere
particolare. Perciò le innovazioni sorgono indipendentemente dalle funzioni per
le quali eventualmente saranno scelte. Agendo come un setaccio, la
selezione naturale può soltanto setacciare differenzialmente ciò che
le viene presentato. Una innovazione deve essersi prodotta necessariamente in
qualche altro modo, come le pepite d'oro separate dal pietrisco. Gli ingredienti
antecedenti al linguaggio devono già esistere in un certo senso. Ma che
cos'erano quegli ingredienti?
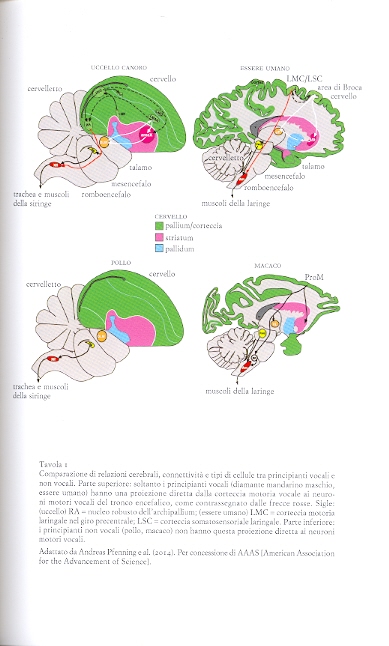
Modello tripartito, apprendimento vocale e genomica
Ogni resoconto sull'origine del linguaggio deve fare i conti con il che cosa si è evoluto. Nel nostro quadro tripartito, opera naturalmente come ciascuno dei tre componenti che abbiamo delineato sopra: 1) l'operatore combinatorio Fondere, insieme con elementi atomici simili a parole, grosso modo il «processore centrale» (CPU [Central Processing Unit] ) della sintassi del linguaggio umano; e le due interfacce, 2) l'interfaccia sensomotoria che fa parte del sistema del linguaggio per l'esternalizzazione, compresi l'apprendimento e la produzione vocale e 3) l'interfaccia concettual-intenzionale, per il pensiero. In questa sede ci concentriamo su (2), sull'apprendimento e la produzione vocali, in quanto mediati dall'interfaccia sensomotoria. [...] Almeno per noi, l'esperimento di Schreiweis insieme con le scoperte di Andreas Pfenning e colleghi (2014) confermano in modo sorprendente che l'apprendimento e la produzione vocale del sistema di esternalizzazione del linguaggio non sono un aspetto specificamente umano. Un tempo pari all'incirca a 600 milioni di anni di evoluzione ci separa dagli uccelli; eppure le regioni specializzate del canto e della parola e la specializzazione genomica dell'apprendimento vocale delle specie di uccelli canori (per esempio il diamante mandarino, il colibrì) e quelle dell'apprendimento vocale della specie umana sono a quanto pare marcatamente simili e convergenti. Per contro, i principianti aviari non vocali (polli, quaglie, piccioni) e i principianti non umani non vocali (macachi) non condividono queste specializzazioni genomiche con i principianti vocali (uccelli canori o esseri umani). [...] Sin dall'analisi pionieristica di Marie-Claire King e Alan Wilson (1975), sappiamo che gli esseri umani e gli scimpanzé sono per il 99% identici a livello macromolecolare (proteine coinvolte nel funzionamento della biochimica degli organismi) e che questa identità risulterebbe probabilmente ancora più stretta, se confrontassimo gli esseri umani con i nostri antenati non umani. King e Wilson traevano questa conclusione ovvia e importante: le differenze tra gli esseri umani e gli scimpanzé devono risiedere in gran parte negli elementi regolatori. Significa che i cambiamenti dei geni che codificano le proteine potrebbero non essere la sede dell'azione evolutiva, in particolare - forse - nell'evoluzione che ci ha fatto esseri umani, perché si è trattato di un evento relativamente recente. Nel corso degli ultimi quarant'anni l'importante intuizione di King e Wilson è stata incontestabilmente confermata, inclusi sia il DNA non codificante, sia tutti gli altri componenti che regolano l'attività dei geni, dall'impalcatura di cromatina che circonda il DNA, alla regolazione microRNA del DNA durante lo sviluppo, in particolare nello sviluppo del cervello: si tratta di una parte della cosiddetta rivoluzione «evo-devo» (Somel, Liu e Khaitovich 2013). | << | < | > | >> |Pagina 49Che altro può dirci la moderna biologia molecolare circa l'evoluzione del cervello e del linguaggio umani? In questa sede non possiamo rendere giustizia a questo campo in rapida espansione, ma possiamo isolare alcuni punti fondamentali insieme con ben noti grandi «blocchi stradali».Anzitutto, grazie al recente lavoro con DNA antico, adesso è possibile riuscire a immaginare quante e che tipo di differenze ci possiamo aspettare di scoprire, per poi vedere come tutto ciò si allinea con le note differenze genomiche tra noi e i genomi sequenziati dell'uomo di Neanderthal, di Denisova e degli scimpanzé. Quanto alle differenze attese, il periodo di tempo trascorso dal distacco dai nostri antenati estinti Homo, come l'uomo di Neanderthal, è relativamente recente (da 500 a 700 000 anni fa), e gli esseri umani moderni sono comparsi nell'Africa meridionale circa 200 000 anni fa, per cui ci sono circa 400 000 anni di tempo evolutivo tra i due eventi. Possiamo impiegare strumenti teorici di genetica delle popolazioni, includendo le stime della forza selettiva, l'ampiezza della popolazione e i tassi di mutazione del DNA, per calcolare quante regioni genomiche distinte, positivamente selezionate, ci possiamo aspettare di scoprire fisse nella popolazione umana (cioè senza variazioni negli esseri umani moderni, e quindi presumibilmente importanti da un punto di vista funzionale), ma differenti nelle specie non umane. La cosiddetta ampiezza della popolazione effettiva degli esseri umani di 200 000 anni fa è stata stimata da diverse fonti pari a circa 10 000 unità, quindi relativamente piccola se confrontata con molti altri mammiferi (Jobling e al. 2013). La forza selettiva (denotata con s ) è difficile da stimare in qualsiasi situazione, ma possiamo usare i dati desunti da uno dei più forti segnali recenti della selezione nella popolazione, cioè quello del gene LCT della persistenza della lattasi (Tishkoff e al. 2006), per dare un limite superiore di 0,10. Questo limite è estremamente elevato. Dati tutti questi parametri, un'analisi recente stima che ci sarebbero state 700 mutazioni benefiche, di cui soltanto 14 sono sopravvissute fissandosi nella popolazione umana, anche dato un forte vantaggio selettivo di s=0,01 (Somel, Liu e Khaitovich 2013). La bassa sopravvivenza è dovuta all'effetto «pozzo gravitazionale stocastico» descritto nel paragrafo precedente, con una probabilità di perdita approssimativamente pari a (1 - s/2): quindi il 98% di 700, ossia 686 perse e 14 fisse. Questa stima teorica risulta abbastanza vicina a ciò che è stato scoperto empiricamente. Il sequenziamento dell'intero genoma dei Neanderthaliani e dei Denisoviani indica che esistono rispettivamente 87 e 260 differenze genomiche funzionali (cambio di amminoacidi), le quali sono fissate negli esseri umani moderni, ma non presenti in queste due specie estinte (Pääbo 2014a, tavola supplementare 1). | << | < | > | >> |Pagina 53Qual è la conclusione? Forse la domanda da un milione di dollari è se i Neanderthaliani avevano il linguaggio. Il numero delle differenze genomiche tra noi e i Neanderthaliani e i Denisoviani è così piccolo che alcuni autori rispondono di sì. Noi continuiamo a essere scettici. Non comprendiamo la base genomica o neurale della proprietà fondamentale. È virtualmente impossibile persino dire se gli esseri umani anatomicamente moderni avessero il linguaggio 80 000 anni fa. Possiamo soltanto affidarci ai sostituti simbolici del comportamento linguistico. Osserviamo con Tattersall (2010) che le prove materiali del comportamento simbolico neanderthaliano sono straordinariamente scarne. Per contro, gli esseri umani anatomicamente moderni nell'Africa meridionale intorno a 80 000 anni fa dimostrano chiari segni di un comportamento simbolico, prima del loro esodo in Europa. Nel capitolo 4 ritorneremo sulla questione.Il problema generale è la nostra scarsa comprensione persino del modo in cui le operazioni computazionali più elementari possono essere effettuate nel sistema nervoso centrale [neural «wetware»]. | << | < | > | >> |Pagina 84Se il quadro delineato grosso modo tiene, può ottenere una risposta la seconda delle due domande fondamentali poste all'inizio di questo capitolo: perché esistono così tante lingue? La ragione potrebbe essere che il problema dell'esternalizzazione è risolvibile in molti modi diversi e indipendenti, prima o dopo la dispersione della popolazione originaria. Non abbiamo motivo di supporre che per risolvere il problema dell'esternalizzazione sia richiesto un cambiamento evolutivo, cioè un cambiamento genomico. Potrebbe essere semplicemente un problema affrontato da processi cognitivi già esistenti, in modi e in tempi diversi. Esiste talvolta una tendenza malaugurata a confondere il cambiamento evolutivo (genomico) in senso letterale con il cambiamento storico: due fenomeni del tutto distinti. Come abbiamo già osservato, esistono prove evidentissime del fatto che non ci fu alcuna evoluzione di rilievo della facoltà del linguaggio dopo l'esodo dall'Africa circa 60 000 anni fa, sebbene sia indubbio che ci siano stati grandi cambiamenti, e anche invenzioni di modi di esternalizzazione (come nel linguaggio dei segni). Si eviterebbe la confusione su questi argomenti sostituendo le nozioni metaforiche «evoluzione del linguaggio» e «cambiamento del linguaggio» con le loro corrispondenti più esatte: evoluzione degli organismi che usano il linguaggio e cambiamento dei modi in cui lo fanno. Detto così in maniera più precisa: l'emergere della facoltà del linguaggio comportò evoluzione, mentre il cambiamento storico (che continua costantemente) non la comportò.Ancora una volta, questi assunti sembrano i più semplici, e non si conoscono ragioni per respingerli. Se in linea di massima sono sulla strada giusta, ne consegue che l'esternalizzazione può non essersi evoluta affatto. Invece avrebbe potuto essere un processo di problem solving, di soluzione di un problema con l'impiego di capacità cognitive già presenti in altri animali. L'evoluzione nel senso biologico del termine sarebbe allora limitata ai cambiamenti che produssero Fondere e la Proprietà Fondamentale, accanto a qualunque altro elemento residuo resista a una spiegazione in termini di Tesi Minimalista Forte, e a qualunque vincolo specifico del linguaggio possa essere imposto alla soluzione del problema cognitivo dell'esternalizzazione. Perciò qualunque modo di affrontare l'«evoluzione del linguaggio» che si concentri sulla comunicazione, o sul sistema sensomotorio, o sulle proprietà statistiche del linguaggio parlato, e via dicendo, potrebbe essere gravemente fuorviante. Questo giudizio copre un ambito piuttosto vasto, come comprenderà chi conosce la letteratura in materia. | << | < | > | >> |Pagina 87Le capacità conoscitive degli esseri umani ci forniscono un mondo di esperienze differente dal mondo di esperienze degli altri animali. Essendo creature riflessive, grazie all'emergere della capacità umana, gli esseri umani cercano di attribuire un senso all'esperienza. Questi tentativi si chiamano «miti», «religione», «magia», «filosofia» o, nell'uso inglese moderno, «scienza». Per la scienza, il concetto di riferimento in senso tecnico è un ideale normativo: speriamo che i concetti inventati «fotone» o «sintagma verbale» colgano una cosa reale nel mondo. E naturalmente il concetto di riferimento va bene per il contesto per il quale è stato coniato nella logica moderna: nei sistemi formali, nei quali si stipula la relazione di riferimento, che vige per esempio tra i numerali e i numeri. Ma il linguaggio e il pensiero umani non sembrano funzionare così, ed equivoci senza fine sono nati dal mancato riconoscimento di questo fatto.Qui tocchiamo argomenti vasti ed estremamente interessanti, che dobbiamo accantonare. Riassumiamo solo brevemente quella che sembra essere, al momento, la migliore ipotesi circa l'unità e la diversità del linguaggio e del pensiero. In maniera del tutto sconosciuta i nostri antenati svilupparono concetti umani. A un certo punto del passato recentissimo, a quanto pare un po' prima di 80 000 anni fa, considerando gli annessi sostituti simbolici, alcuni individui di un ristretto gruppo di ominidi dell'Africa orientale subirono un piccolo cambiamento biologico che consentì l'operazione Fondere: un'operazione che assume i concetti umani come atomi computazionali e produce espressioni strutturate che, interpretate sistematicamente dal sistema concettuale, forniscono il ricco linguaggio del pensiero. Questi processi potrebbero essere computazionalmente perfetti, o quasi, e quindi risultanti da leggi fisiche indipendenti dagli esseri umani. L'innovazione ebbe ovvi vantaggi e si impose sull'intero gruppo. In una fase successiva il linguaggio interno del pensiero venne collegato al sistema sensomotorio: un compito complesso che può essere risolto in molti modi differenti e in diversi periodi di tempo. Nel corso di questi eventi la capacità umana prese forma, producendo buona parte della nostra «natura intellettuale e morale», secondo l'espressione di Wallace. Gli esiti sembrano essere molto diversi, però hanno un'unità sostanziale, che riflette il fatto che gli esseri umani sono identici per gli aspetti fondamentali, proprio come l'ipotetico scienziato extraterrestre che abbiamo evocato in precedenza potrebbe concludere che esiste soltanto una lingua con piccole variazioni dialettali, principalmente (o forse interamente) nel modo dell'esternalizzazione. In conclusione, ricordiamo che, anche se questa storia nelle sue linee generali ha grosso modo una tenuta, e le enormi lacune possono essere colmate, rimarranno sempre problemi irrisolti, sollevati da centinaia di anni. Tra questi c'è la questione relativa al modo in cui le proprietà definite «mentali» sono in relazione con «la struttura organica del cervello», secondo la formulazione del XVIII secolo, e ci sono i problemi più misteriosi dell'uso ordinario, creativo e coerente, del linguaggio, che costituiva l'interesse principale della scienza cartesiana e che senza dubbio non è ancora all'orizzonte dell'indagine. | << | < | > | >> |Pagina 893.
L'architettura del linguaggio e la sua importanza per l'evoluzione
L'indagine razionale sull'evoluzione di un sistema può procedere evidentemente solo nella misura in cui se ne comprenda la natura. In maniera non meno evidente, senza una seria comprensione della natura fondamentale di un sistema, le sue manifestazioni appariranno caotiche, estremamente variabili e prive di proprietà generali. Di conseguenza, lo studio della sua evoluzione non potrà essere intrapreso seriamente. Anche questa indagine dev'essere il più possibile fedele a tutto ciò che sappiamo della storia evolutiva. Verità lapalissiane che valgono per lo studio della facoltà del linguaggio umano, così come per altri sistemi biologici. Le proposte formulate nella letteratura in materia si possono valutare sulla base della loro aderenza a tali condizioni elementari. Il problema dell'evoluzione del linguaggio sorse a metà del XX secolo con i primi tentativi di costruire resoconti del linguaggio come oggetto biologico, interno a un individuo, e di catturare quella che possiamo chiamare «Proprietà Fondamentale» del linguaggio umano: ciascuna lingua produce un assortimento digitalmente infinito di espressioni strutturate gerarchicamente con interpretazioni sistematiche all'interfaccia con altri due sistemi interni: il sistema sensomotorio per l'esternalizzazione e il sistema concettuale per l'inferenza, l'interpretazione, la progettazione, l'organizzazione dell'azione, e per altri elementi di quello che informalmente è chiamato «pensiero». L'impostazione generale che per il linguaggio adotta queste linee-guida è stata chiamata «programma biolinguistico». Nella terminologia corrente, una lingua intesa in questi termini è chiamata «lingua interna» o «I-lingua». In virtù della Proprietà Fondamentale, ciascuna I-lingua è un sistema di «segni udibili per il pensiero», per citare il grande indoeuropeista William Whitney (1875, p. 3), anche se adesso sappiamo che non è necessario delimitare l'esternalizzazione alle modalità articolatorio-uditive. Per definizione, la teoria di una I-lingua è la sua grammatica generativa, e la teoria generale delle I-lingue è la Grammatica Universale (GU), adattando nozioni tradizionali a un contesto nuovo. GU è la teoria del componente genetico della facoltà del linguaggio, la capacità che rende possibile l'acquisizione e l'uso di I-lingue particolari. GU determina la classe dei procedimenti generativi che soddisfano la Proprietà Fondamentale e gli elementi atomici che intervengono nei computi. | << | < | > | >> |Pagina 1064.
Triangoli nel cervello
Fuori della portata della selezione naturale?
Alfred Wallace
Così Wallace si diede al crimine: il crimine di spostare la selezione fuori della portata della selezione «naturale»: «Perciò dobbiamo ammettere la possibilità che nello sviluppo della razza umana un'intelligenza superiore abbia guidato le stesse leggi [della variazione, della moltiplicazione e della sopravvivenza] per fini più nobili» (1869, p. 394). Darwin rimase esterrefatto. Scrisse a Wallace: «Mi auguro che tu non abbia massacrato del tutto il bambino, tuo e mio» (Marchant 1916, p. 240). Secondo noi il «crimine» di Wallace non fu, in fondo, un peccato grave. Egli aveva semplicemente messo in luce la verità: il darwinismo prevedeva una continuità strettamente graduale con il passato, «numerose, successive, lievi modificazioni» tra i nostri antenati e noi. Eppure esiste un abisso spalancato tra ciò che noi possiamo fare e ciò che gli altri animali non possono fare: il linguaggio. E qui c'è un mistero. Come con qualsiasi autentico mistero, dobbiamo figurarci un «giallo»: che cosa, chi, dove, quando, come e perché. In questo capitolo faremo del nostro meglio per rispondere a ciascuna di queste domande. In sintesi, le nostre risposte alle domande sul linguaggio avranno l'andamento seguente: - il «che cosa» si riduce alla Proprietà Fondamentale del linguaggio umano: la capacità di costruire una gamma digitalmente infinita di espressioni strutturate gerarchicamente con determinate interpretazioni all'interfaccia con altri sistemi organici; - il «chi» siamo noi (esseri umani anatomicamente moderni), e non gli scimpanzé, né i gorilla, né gli uccelli canori; - il «dove» e il «quando» indicano un lasso di tempo collocabile tra la prima comparsa di esseri umani anatomicamente moderni in Africa meridionale, circa 200 000 anni fa, e l'ultimo esodo dall'Africa, circa 60 000 anni fa (Pagani e al. 2015); - il «come» è l'implementazione neurale della Proprietà Fondamentale, di cui sappiamo ben poco, ma le prove empiriche recenti indicano che la cosa potrebbe essere compatibile con un «leggero ricablaggio del cervello», come abbiamo detto altrove; - il «perché» è l'uso del linguaggio per il pensiero interno, come collante cognitivo che lega insieme altri sistemi cognitivi deputati alla percezione e all'elaborazione dell'informazione. Da quanto riusciamo a capire, questo quadro dell'evoluzione del linguaggio umano si inserisce in maniera eccellente nella concezione di Jacob e Monod dell'evoluzione per selezione naturale come bricolage opportunistico. Noi sosteniamo che gran parte degli ingredienti necessari per il linguaggio umano fossero a disposizione già prima. Lo scopo dei circuiti corticali esistenti venne ridefinito. Poi certi piccoli cambiamenti genomici determinarono effetti cognitivi relativamente estesi: si tratta precisamente del quadro delineato da Ramus e Fisher (2009) che abbiamo citato nel capitolo 2. A differenza di altri, non pensiamo che ci sia bisogno di chiamare in causa «chiacchiere», una versione pleistocenica di «Google maps», o un'evoluzione culturale di tipo misterioso. | << | < | > | >> |Pagina 135Chi?Sappiamo che gli animali non umani eccellono in molti compiti cognitivi impegnativi. I corvidi (famiglia a cui appartengono i corvi) sono molto intelligenti in svariati domini cognitivi. Hanno la capacità di fabbricare strumenti, di effettuare ragionamenti spaziali e causali sofisticati, e ricordano la localizzazione e la qualità del cibo nascosto. Le ghiandaie occidentali sono in grado di calare fili legati attorno a piccoli ciottoli nelle fessure per adescare le formiche per il pasto. Sebbene alcuni uccelli come le quaglie e i polli non siano principianti vocali, gli uccelli canori sono capaci di imparare vocalizzazioni molto sofisticate. Gli uccelli canori maschi istruiscono i piccoli, che devono imparare il canto dell'istruttore, talvolta con piccole modifiche, così da poter usare i loro canti come richiami territoriali o di disponibilità sessuale. Come negli esseri umani, c'è anche una lateralizzazione cerebrale sinistra. E come negli esseri umani, esiste un periodo critico per l'apprendimento, interrotto alla pubertà dal testosterone. Con somiglianze così numerose ed evidenti, non c'è da meravigliarsi che ai tempi di Aristotele ci si sia chiesti se i canti degli uccelli costituissero un buon modello per il linguaggio. Tuttavia, stando a quello che sappiamo oggi, possiamo concludere che il canto degli uccelli è soltanto un modello per la lingua parlata, caso mai, ma non per il linguaggio. Come Berwick e colleghi (2011, p. 114) hanno rimarcato, «la maggior parte delle proprietà sintattiche del linguaggio umano non si trova nel canto degli uccelli. L'unica eccezione si riferisce alle proprietà dei sistemi fonici del linguaggio umano». La cosa è abbastanza evidente in qualunque analisi formale del canto degli uccelli e nelle sue somiglianze e differenze rispetto al linguaggio umano (un sommario nella tabella 1, ibid.). Delle sedici proprietà fondamentali della sintassi del linguaggio umano, soltanto due si trovano nella sintassi sia del canto degli uccelli sia del linguaggio umano: dipendenze basate sull'adiacenza e raggruppamento in «porzioni».
Il tutto è riassumibile così: sia il canto degli uccelli sia il sistema fonico per l'esternalizzazione delle lingue umane hanno dipendenze basate sulla precedenza, descrivibili attraverso reti di transizione a stati finiti. Tutte le altre proprietà fondamentali della sintassi del linguaggio umano non sono presenti nel canto degli uccelli. Queste proprietà comprendono le dipendenze illimitate non adiacenti, la struttura gerarchica, la dipendenza delle regole sintattiche dalla struttura e l'apparente «dislocazione» di sintagmi con copie. [...] Data questa «disposizione linguistica» all'apprendimento e alla produzione vocali, se il cervello dei primati è davvero «sintonizzato» con le proprietà fonetiche o fonemiche del linguaggio, ma le scimmie non sentono che rumore mentre i bambini estraggono dal rumore un materiale linguisticamente pertinente, abbiamo subito una prova a favore di un certo tipo di elaborazione interna, unico per i neonati umani e assente in altri primati. Per adesso accantoniamo il rompicapo. [...] Che dire degli altri animali non umani? I nostri più stretti parenti vivi, i primati non umani, sono stati indicati a lungo come buoni candidati. Ci si stupirà, ma risulta che abbiano le stesse limitazioni degli uccelli canori. Per esempio, sono stati fatti parecchi famosi tentativi di «insegnare» agli scimpanzé il linguaggio umano. Il Progetto Nim è tra i più noti. I ricercatori della Columbia University avevano tentato di insegnare a Nim la ASL [ American Sign Language, la lingua dei segni americana]. Non ci riuscirono. Ciò che Nim fu capace di imparare dell'ASL fu una specie di memorizzazione meccanica: sequenze di segni lineari (corte). Non progredì mai fino a produrre frasi incassate, chiaramente strutturate gerachicamente, come ogni bambino normale riesce a fare all'età di tre o quattro anni. (Vedremo tra breve in che modo siamo in grado di determinarlo formalmente). Se Nim voleva una mela, scorreva il catalogo di tutti i singoli segni associati a mela, recuperando Nim mela, mela Nim, mela coltello e così via: come dice Laura-Ann Petitto (2005, p. 85), una neuroscienziata che lo seguiva, «costruiva una lista della spesa» delle «parole» che conosceva meglio. Nim non raggiunse il livello di capacità sintattica di un bambino di tre anni; nessuna struttura gerarchica trovò conferma. [...] Se ci riflettiamo un momento, è evidente che gli scimpanzé sono esempi perfetti di «principianti associazionisti» puri: a quanto pare, hanno connessioni immediate tra particolari stimoli esterni e i loro segni. Non sembrano considerare la mela che vedono in un qualche modo dipendente dalla mente, come è stato detto nel capitolo 3. Invece hanno immagazzinato una lista di associazioni esplicite, indipendenti dalla mente, tra oggetti del mondo esterno e i loro segni ASL. Siamo ben lontani dalla capacità linguistica di tipo umano: gli scimpanzé non hanno né Fondere né gli elementi simili alle parole di cui sono in possesso gli uomini. Se è così, anche gli scimpanzé sono eliminati come sospetti nel nostro giallo. | << | < | > | >> |Pagina 144Dove e quando?Se la Proprietà Fondamentale è veramente fondamentale, dove e quando è comparsa per la prima volta? Come abbiamo osservato nel capitolo 3, è opinione comune che l'origine degli elementi dipendenti dalla mente e simili alle parole rimanga un bel mistero: per chiunque, noi compresi. [...] Se facciamo affidamento sulle prove univoche del comportamento simbolico come surrogato del linguaggio, potremmo assumere che i manufatti della grotta di Blombos nell'Africa meridionale (pezzi di ocra incisi con disegni geometrici e perline) forniscano la più ragionevole indicazione spazio-temporale per la comparsa del linguaggio, cioè circa 80 000 anni fa, proprio in quel punto. Come si è detto nel capitolo 1, pare esista un ampio «scollegamento» tra la comparsa di cambiamenti morfologici nella morfologia di Homo e qualunque cambiamento comportamentale o tecnico associato: la comparsa di nuove tecniche e di nuovi comportamenti segue a lunghi periodi di stasi dopo la comparsa di nuove varianti di Homo. Cosicché possiamo fissare il «quando» tra due punti temporali: tra la comparsa di esseri umani anatomicamente moderni, all'incirca 200 000 anni fa nell'Africa meridionale, e i primi esseri umani moderni sotto il profilo comportamentale, grosso modo 80 000 anni fa. Poi, all'incirca 60 000 anni fa, arrivò l'esodo africano, con esseri umani pienamente moderni che si espansero nel Vecchio Mondo e in seguito in Australia. L'assenza di qualsiasi variazione profonda nella facoltà del linguaggio indica la stessa conclusione. C'è qualche ragione di dubitare che un bambino di una tribù di Papua Nuova Guinea, rimasta per 60 000 anni senza contatti con altri gruppi umani, crescendo a Boston fin dalla nascita sarebbe diverso da un bambino del luogo? Nessuna che siamo in grado di stabilire. L'aneddoto raccontato da Ledyard Stebbins a proposito di Theodosius Dobzhansky ed Ernst Mayr (cfr. supra, p. 58, nota 1) è una versione moderna dell'esperimento, e anche lavori recenti di genomica giungono alle stesse conclusioni)' Quindi, a nostro avviso, il linguaggio umano e la Proprietà Fondamentale devono essere sorti tra questi due estremi temporali, 200 000 anni fa al più presto e 60 000 anni fa al più tardi, ma presumibilmente ben prima dell'esodo africano, date le testimonianze simboliche della grotta di Blombos, risalenti a 80 000 anni fa. Certo, se emergessero altre prove, la data potrebbe essere anticipata, e magari avvicinata di più alla tacca dei 200 000 anni. [...]
Riassumendo, la stima migliore sul «quando» e sul «dove» è un
certo momento collocabile tra la comparsa dei primi esseri umani
anatomicamente moderni nell'Africa meridionale, circa 200 000
anni fa, e il periodo precedente l'ultimo esodo dall'Africa, circa
60 000 anni fa (Pagani e al. 2015), ma probabilmente prima di
80 000 anni fa. Circa 130 000 anni quindi, un lasso di tempo per il
cambiamento evolutivo pari grosso modo a 5000-6000 generazioni.
Non «da giorno all'altro in una generazione», come alcuni hanno
(erroneamente) inferito, ma nemmeno un periodo misurabile su
una scala di eoni geologici. Un lasso di tempo sufficiente, all'interno
dell'approssimazione stimata da Dan Nilsson e Susanne Pelger (1994) come il
periodo necessario per l'evoluzione completa di un occhio di vertebrato a
partire da una singola cellula, anche senza invocare effetti «evo-devo».
|